
bplant.org Blog
How To Identify Plants
May 7th, 2026 by Alex Zorach
One of the questions I get asked most frequently is: how do you identify plants? There is no short answer to this question, other than with great effort. But I hope this post can describe the process I use so that it might help others. The most important thing about plant ID is to understand that it is a lifelong process and you will have the best results if you approach it both with a zeal for observing and learning, and with humility, realizing that you will always know little relative to the totality of what there is to know. What is this plant, which was observed growing in a moist woodland opening in Mammoth Cave National Park, in Edmonson County, KY? How would you figure out what it is? Click here for a picture of its habitat and growth habit. Try reading this blog post and testing your newfound skills, then Click here for the answer. Public Domain photo by Clare Bledsoe, Source.
What is this plant, which was observed growing in a moist woodland opening in Mammoth Cave National Park, in Edmonson County, KY? How would you figure out what it is? Click here for a picture of its habitat and growth habit. Try reading this blog post and testing your newfound skills, then Click here for the answer. Public Domain photo by Clare Bledsoe, Source.{kind=link}
But there are also numerous tips and tricks, each of which can lead to small breakthroughs. My goal with this post is to collect all of my insights into one place so that someone reading it can dramatically ramp up their ability to ID plants, or at least, become much faster and more efficient at learning how to ID plants.
Learning from Birdwatchers
One of the best insights I ever had about how to identify plants came from a book about identifying not plants, but birds: The Field Guide to Advanced Birding by Kenn Kaufman. Unlike most field guides, which are picture-rich depictions of what each species looks like, this prose-rich meta guide explains how to look at birds and how to get better at looking at birds. Pine warbler (Setophaga pinus) in fall plumage is often considered one of the "confusing fall warblers" which are notoriously hard to ID. A lot of plants can be harder to ID than even these warblers, due to the massive number of species. But, unlike birds, plants sit still. Photo © Laura Clark, CC BY 4.0, Source.
Pine warbler (Setophaga pinus) in fall plumage is often considered one of the "confusing fall warblers" which are notoriously hard to ID. A lot of plants can be harder to ID than even these warblers, due to the massive number of species. But, unlike birds, plants sit still. Photo © Laura Clark, CC BY 4.0, Source.Kaufman notes that a lot of beginner birdwatchers look at a bird only until they are sure what it is, and then they immediately turn their attention to some other bird that they have not yet identified. His core advice is to spend more time observing birds that you have already identified, starting with the most common species where you already know exactly what they are. This advice humbled me as I had been doing exactly what he was telling me not to do.
By looking at a bird whose identity you already know, you will learn that species better. But you also hone your general skills for looking closely at birds. This ensures that when do do see something new and different, you will immediately know that it is something unfamiliar, and you will also be better able to articulate how it is different and how you recognized it as different.
This same process can apply to plants. So I recast his advice to the world of plants: if you want to get better at identifying plants, start by looking at the plants whose identity you already know. And don't forget how lucky you are, compared to birdwatchers: the plants sit still and you can take your time and get as close as you want to.
Why I Treat Keys With Caution
When I first started studying plant identification I focused on dichotomous keys, those long decision trees asking question after question. After years of wrestling with these keys, I concluded that in a majority of the cases and for most people, keys are not the best way to identify plants. Even when they work, they need to be balanced with and checked against other methods. This screenshot shows part of the Carex Key from Flora of North America. This key is divided into 6 sections and this section has 33 entries. The key is so large both because the Carex genus is diverse, and because FNA covers all of North America. It requires access to the plant at the time of blooming and/or seed production, and knowledge of specialized botanical terminology. The decisions in the key do not correspond to taxonomic relationships, and some taxa appear in the key on multiple lines.
This screenshot shows part of the Carex Key from Flora of North America. This key is divided into 6 sections and this section has 33 entries. The key is so large both because the Carex genus is diverse, and because FNA covers all of North America. It requires access to the plant at the time of blooming and/or seed production, and knowledge of specialized botanical terminology. The decisions in the key do not correspond to taxonomic relationships, and some taxa appear in the key on multiple lines.What can go wrong with keys?
- Keys waste time by having you check through plants that, for whatever reason, are not an option in your particular scenario. The effort necessary to accurately navigate each choice in a key is usually greater than the effort necessary to exclude most of the other options. For example, you might be using a key for Pennsylvania which checks against all plants possible anywhere in the state, and many of them may not occur in your region.
- Keys are not robust with respect to errors. In order to get a correct identification from a key, every choice must be correct. Sometimes when you make an error, it is not apparent until you get to the end of the process, and then you need to find the error, which can be time-consuming.
- The choices in keys do not always correspond to taxonomic relationships (which taxa are closest-related to each other), so time invested in using keys may not help learn these relationships.
- Keys can be wrong, especially in the case of aberrant individual plants, which are more common than you might realize. Dimensions, counts, color, texture, and other aspects of plant morphology, as well as bloom timing, can all be outside the parameters of the descriptions used in keys on some individuals, both due to natural variation and stressors on the plant.
- Keys often reference aspects of a plant that are not visible at all times of year, such as flowers or seeds, even in cases where the plants can be clearly identified through other features such as leaves, habitat, or range.
- Keys often require more knowledge of botanical terminology than is necessary to confidently identify a species. The terminology is necessary not for the keys to function, but for them to be concise enough that the volumes containing the keys do not end up reaching an unmanageable length, and so that they can be practically written and maintained by their authors or maintainers.
 The populations of tall blue lettuce (Lactuca biennis) in and around northern Delaware, including into southeastern PA, have a rare pale yellow color morph. Many keys for Lactuca assume L. biennis flowers are pale blue to white, which can lead to confusion. Note that FSUS's key for Lactuca acknowledges the flowers of this species can be "rarely yellow". In my experience FSUS has more reliable keys than most, but even there it is possible to find individual plants for which a key will not yield the correct ID. Photo © Alex Zorach, CC BY 4.0, Source.
The populations of tall blue lettuce (Lactuca biennis) in and around northern Delaware, including into southeastern PA, have a rare pale yellow color morph. Many keys for Lactuca assume L. biennis flowers are pale blue to white, which can lead to confusion. Note that FSUS's key for Lactuca acknowledges the flowers of this species can be "rarely yellow". In my experience FSUS has more reliable keys than most, but even there it is possible to find individual plants for which a key will not yield the correct ID. Photo © Alex Zorach, CC BY 4.0, Source.I am not saying never to use keys. They have their purpose, and as you learn more you will likely get more out of them. But if you are starting out, you can save a lot of time and learn a lot faster through other approaches. Then return to the keys when you know how to interpret them, and use them to rigorously confirm an ID you are already relatively certain of, or perhaps to discover a rare but possible plant that isn't showing up through your other methods.
So if not keys, what to do?
My approach to plant identification is as follows:- Learn common, easy-to-identify plants first.
- Closely observe plants you have already identified.
- Identify to a higher taxon if possible, such as genus, family, or other grouping.
- Check which plants are possible based on range.
- Look at habitat in depth.
- Once you've narrowed it to a reasonable number of species, read descriptions and look up pictures of those species.
- Use tools to check for common lookalikes that might be outside the subset of plants you are examining (I outline a process for this below.)
- If you cannot even get started, use AI-based tools (I will highlight which ones I prefer below) to make an initial guess. Then check the family, genus, species, or other taxon rigorously.
By the time you've done all these things, you will usually know inside and out, which plant you are looking at, and you'll be able to identify how and why you know. But, more importantly, all of the extra work you are doing will help prepare you to later identify other plants. Plant ID is less about identifying individual plants in isolation, and more about building a web of knowledge. It may seem time-consuming to identify a single plant, but you can take solace in the fact that in the course of doing so, you lay the foundations to identify many other plants more easily in the future.
I will now go through each of these points in detail to show you how I do them.
Learn Common, Easy-to-Identify Plants First
Some plants are easier to identify than others. For example, in most of Eastern North America, American pokeweed (Phytolacca americana) is not only abundant, but nothing else looks remotely like it (three other Phytolacca species do occur in North America but all are exceedingly rare.) Ginkgo (Ginkgo biloba) has a visually-distinctive leaf. Because it is easy to definitively ID, it can be a good starting point for the process of looking more closely at plants. Once you have identified a tree as a gingko, you can begin looking at more subtle aspects of its morphology, such as its bark or form. Photo © Sandy Wolkenberg, CC BY 4.0, Source.
Ginkgo (Ginkgo biloba) has a visually-distinctive leaf. Because it is easy to definitively ID, it can be a good starting point for the process of looking more closely at plants. Once you have identified a tree as a gingko, you can begin looking at more subtle aspects of its morphology, such as its bark or form. Photo © Sandy Wolkenberg, CC BY 4.0, Source.From there, you can branch out to learning other plants that are either easy-to-identify or common, but not both. For example, Gingko is occasional in landscaping and a rare escape in the wild, but is visually-distinctive and easy to identify, so you can learn to ID it with minimal effort. Oaks (Quercus) on the other hand, are harder to ID, but common, so it might be worth learning at least the most common oaks in your region. Even if more effort is required, the fact that they are common will make it easier to learn them and more worthwhile to do so.
Over time, you can advance to learning progressively rarer and more difficult-to-ID plants. These goals are often more attainable if you master a particular taxon in your region, progressing through the more common members of it and then learning the rarest ones last.
Closely observe plants you have already identified.
Once you have successfully identified a plant, now is your time to learn it. Observe the plant through its whole life-cycle. Many plants are easy to identify at a specific time of year, due to a distinctive flower or fruit, but can be hard to identify at other times of year. Watching a plant year-round will expand your ability to identify it at times of year when flowers and/or fruit are absent. For example, just about anyone can identify a fruiting pokeweed plant, but can you identify a pokeweed seedling? How about emerging pokeweed shoots from an already-established plant? How about the dead stalks of pokeweed in late winter? Would you recognize these stems and empty seedheads as belonging to yellow crownbeard (Verbesina occidentalis)? This growth from the previous season has persisted into March of the next year. Identifying plants in a state like pictured here is one of the key things gained by observing a plant year-round. Photo © Becky Dill, Public Domain, Source.
Would you recognize these stems and empty seedheads as belonging to yellow crownbeard (Verbesina occidentalis)? This growth from the previous season has persisted into March of the next year. Identifying plants in a state like pictured here is one of the key things gained by observing a plant year-round. Photo © Becky Dill, Public Domain, Source.By following this process, you will not only expand your ability to learn how to identify a particular plant, but you will be building the knowledge and neural machinery to identify other plants in the future. For example, yellow crownbeard, pictured above, is easily confused with wingstem (Verbesina alternifolia). If you've gotten to know yellow crownbeard, and you come across an individual of wingstem, even if you have never seen it before, you may recognize it as something related but not exactly the same. This realization could prompt you to examine other species in the same genus, which in this case would lead you to a correct ID.
Deciduous woody plants are usually easiest to identify when leaves and/or flowers are present. Once you have identified one, you can then watch it through the winter (or dry season in an area where plants are drought-deciduous) and learn to identify it by aspects like its bark, form, branching habit, or buds. Sometimes these other aspects of a plant are more distinct than their leaves. For example hackberries (Celtis sp.) have leaves and a growth habit that resembles elms (Ulmus sp.), but their warty bark is entirely unlike that of elms.
Identify to a Higher Taxon if Possible
Understanding how taxonomy works and knowing some of the more common groupings into which plants are classified can make plant ID a lot easier. Like other living organisms, plants are classified in a hierarchical organization with layers or levels of names, called ranks. Scientific names take the form Genus species, and there are also lower (more specific) ranks such as subspecies, variety, and form, and higher (broader or more general) ranks such as family and order. There are also numerous intermediate layers:Kingdom (All Plants) > Division (Phylum) > Class > Order > Family > Subfamily > Tribe > Subtribe > Genus > Section > Species > Subspecies > Variety > Form
Note also that some of the key taxa of plants lie in unnamed ranks between these named ones. For example, the vascular plants (tracheophytes), the flowering plants (Angiospermae), and the gymnosperms are all clades of plants higher than division but lower than class.
The classification of plants in higher taxa than species is subjective. Some genera are large and contain dozens or even hundreds of species (i.e. Astragalus) whereas other genera might contain only one species (i.e. snowplant (Sarcodes sanguinea)) and the same can be true of higher taxa such as families. Similarly, some genera contain plants that look wildly different, such as the bunchberry dogwood (Cornus canadensis), a low-growing herbaceous perennial with a single whorl of leaves, in the same genus as woody plants such as the red-osier dogwood (Cornus sericea). In other cases, a large family might all look similar, such as the Amaryllidaceae family, which is almost all bulbous perennials with linear leaves.
In some cases, you might look at something and identify it to genus level easily, like you might say: "This tree looks like an oak (Quercus sp.)" whereas in other cases you might have no idea of the genus, but identify it to family level, "This plant has a compound flower, with rays around a central disc of tiny flowers, so it is probably in the composite (Asteraceae) family." or "This plant has a square stem, opposite leaves, and a distinctive aroma, so it seems like it might be in the mint (Lamiaceae) family." or you might go only to a high level, such as: "This looks like a monocot (Monocotyledon)." The more plant families, genera, and other groupings you are familiar with, the easier it will be to make these sorts of assessments.
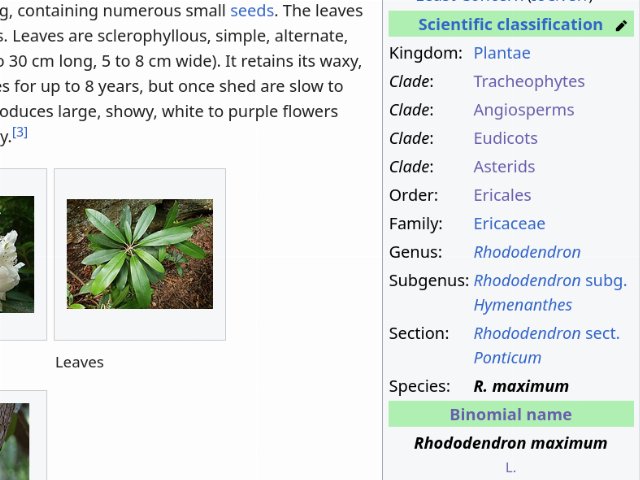 This screenshot shows the taxobox from Wikipedia's page for great laurel (Rhododendron maximum). Note how in this case, there are four clades of unnamed rank between the named rank of Kingdom and Order, and that the named ranks of Division and Class are not listed. Note also that there is a subgenus and section listed. This scenario is typical, although the specifics of how it plays out will vary a lot from one taxon to the next. Also, because Wikipedia is user-edited and worked on incrementally, just because a rank is not listed in the taxobox does not mean that the species is not classified into this rank. Some articles list more detail than others. © Wikipedia, CC BY-SA 4.0, Source.
This screenshot shows the taxobox from Wikipedia's page for great laurel (Rhododendron maximum). Note how in this case, there are four clades of unnamed rank between the named rank of Kingdom and Order, and that the named ranks of Division and Class are not listed. Note also that there is a subgenus and section listed. This scenario is typical, although the specifics of how it plays out will vary a lot from one taxon to the next. Also, because Wikipedia is user-edited and worked on incrementally, just because a rank is not listed in the taxobox does not mean that the species is not classified into this rank. Some articles list more detail than others. © Wikipedia, CC BY-SA 4.0, Source.One of the best tools for learning the taxonomy of various plant species is Wikipedia. Wikipedia has a template called a Taxobox which not only outlines the entire hierarchy of classification, but hyperlinks each higher taxon to the page on that taxon, when it exists. It can be an outstanding way to quickly learn about plant taxonomy. It is, however, a user-generated source in a constant state of flux, so in the case of taxa that have experienced major reclassifications and/or controversy, its information can be incomplete and inconsistent with other sources and sometimes even among its own articles. You can get a lot out of browsing Wikipedia and checking the Taxobox for some of your favorite plants. As you learn which higher taxa each of them is in, you will start seeing patterns, and soon, you will begin to understand how unfamiliar plants fit into these classifications.
For a more rigorous and consistently-updated source of information on plant taxonomy, check Plants of the World Online (POWO) which is run by the Kew Botanic Gardens in London. We use POWO for resolving taxonomy and follow their scheme for most, but not all species. iNaturalist is also an excellent source and has an added advantage of making it easier to browse the taxonomic hierarchy of a particular taxon. iNaturalist, like bplant, usually but not always follows POWO.
Check which plants are possible based on range.
Range is one of the best ways to narrow the possibilities for an unfamiliar plant. However, range is complicated by the fact that plant ranges are not perfectly documented, and can change over time. So, the fact that a plant is not recorded in a particular region does not mean that it could not occur there, just that it is less likely.We recommend using as local sources as possible. Our ecoregion-based range maps are best used only as a starting point. County-level range maps such as those provided by USDA PLANTS and BONAP provide finer detail, but state-level sources such as VA Plant Atlas and Maryland Biodiversity Project are often more up-to-date and sometimes offer still finer detail. FSUS is not quite as fine-tuned but offers data both on range and abundance. Some regions have print sources with verbal descriptions of plant occurences. If you are lucky enough to live in one of these areas, get your hands on these sources! And some parks and nature preserves maintain a plant list, and if you're looking at a plant at one of these sites, that list is a good place to start.
iNaturalist can also be used to narrow down plant ranges, although it is better for common plants and in regions with more data. To use its full features, you will need to access it via the website; the smartphone app supports only some of its features. Its data is user-generated so it is best approached as a suggestion, not rigorous documentation of a plant's presence or absence. The "Explore" feature on iNaturalist can be used to list the species in a particular genus or higher taxon, in a particular location. On the "Explore" page, type any taxon into the "Species" box (it accepts any taxon, not just species), then a region (which could be as fine-tuned as a county or even specific municipality) and then hit the "Species" tab and it will list the species present in observations in that region, in order of frequency of occurrence.
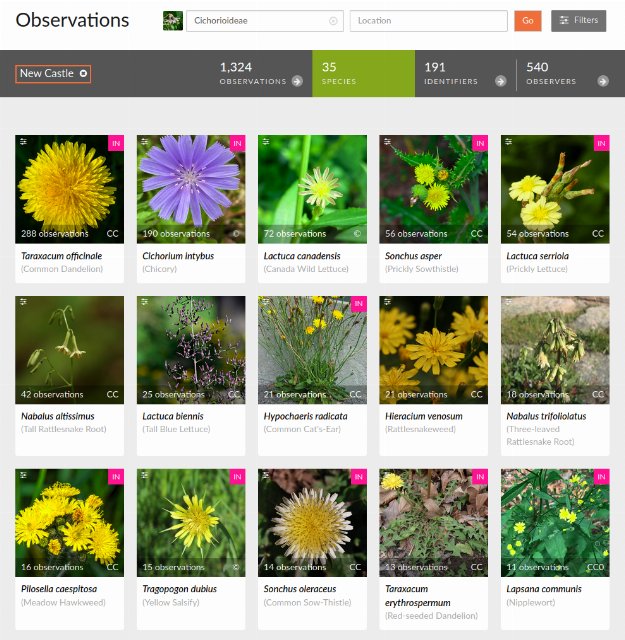 This screenshot shows the species tab on iNaturalist's observation page for the Cichorioideae tribe in New Castle County, Delaware.
This screenshot shows the species tab on iNaturalist's observation page for the Cichorioideae tribe in New Castle County, Delaware.For example, this link shows a list of species in the Cichorioideae tribe of the Asteraceae family, in New Castle County, DE. This list would be good to check against if you see a vaguely dandelion-like flower in this county and have no clue what it is. I live in this county and can testify that the top 5 species not only occur here and are common, but are listed in a reasonable order of how common they are. However, lower down in the list you cannot trust the data to be as useful, as the sample size is too small. I've seen species in Cichorioideae that are not even listed, but the list also contains a number of plants I've never seen here.
Note how this method of narrowing down options by range synergizes with knowledge about higher taxa covered in the previous section. You can combine a region and higher taxon to get a narrower list of likely species than with either factor alone.
Consider out-of-range plants.
Occasionally, you will struggle to ID a plant based on reported ranges, and you must consider the possibility that you have discovered a plant beyond its reported range. Several factors can be useful to assess the likelihood that you have found a plant outside its reported range:- Is the species common or abundant in adjacent areas?
- Is the species increasing or decreasing in the broader region?
- How conspicuous is it?
- How hard is it to ID?
- Is it a species or genus that gets much attention?
It is always best to be cautious and keep considering rare possibilities. When examining reports on iNaturalist of Maryland sanicle (Sanicula marilandica) in unreported counties near the southeastern border of its range, every single one that I was able to identify with confidence turned out to be a misidentification. This species is rare near its southeastern range limits, and is decreasing in these areas, possibly due to global warming. But it ticks all three of the "hard to ID", "inconspicuous", and "doesn't get much attention" boxes, so it's a species I still consider.
Look at habitat in depth.
Habitat is one of the most important ID clues. It is sometimes even easier to observe and/or less variable than the relevant aspects of a plant's shape or appearance necessary to make a definitive ID.For example, the leaf shape of swamp chestnut oak (Quercus michauxii) is similar to both chestnut oak (Quercus montana) and chinquapin oak (Quercus muehlenbergii), and can be easily confused with either species when habitat is not considered. But their habitats are almost entirely non-overlapping: swamp chestnut oak grows in moist bottomlands on acidic soils, whereas chestnut oak grows on dry uplands, also on acidic soils, and chinquapin oak prefers calcareous soils which have higher pH.
Many plants have strict preferences for less-visible aspects of habitat, such as soil pH or nutrient levels. Certain plants only (or never) occur on high pH soils such as rocky limestone or mafic soils, or vice versa on acidic, low pH soils such as occur on coarse sands, highly leached soils, and peaty or mucky soils. Our post thinking more deeply about habitat can help you to learn how to approach habitat more richly, with an eye towards ID.
 This photograph shows a plant of the Eupatorium genus in Westminster, MD. In the Northern Piedmont ecoregion where this plant was photographed, there are 14 Eupatorium species. Using Maryland Biodiversity Project's county-level range maps, this can be narrowed down to 4 possibilities: E. altissimum, perfoliatum, serotinum, and sessilifolium. This individual is Tall thoroughwort (Eupatorium altissimum), a species with a strong preference for higher pH soils and dry, sunny conditions. E. perfoliatum and E. serotinum both prefer moister conditions, and the only other to also occur on dry, higher-pH sites, E. sessifolium, prefers greater shade. The combination of dry, high pH conditions and full sun makes this species the most likely in its genus. Knowing that a site has high soil pH can be as useful as morphology. But conversely, when you have definitively ID'ed a plant as this species, it can function as a clue to the soil conditions, which may help you to identify other plants on the same site. Photo © Tom Field, CC BY 4.0, Source.
This photograph shows a plant of the Eupatorium genus in Westminster, MD. In the Northern Piedmont ecoregion where this plant was photographed, there are 14 Eupatorium species. Using Maryland Biodiversity Project's county-level range maps, this can be narrowed down to 4 possibilities: E. altissimum, perfoliatum, serotinum, and sessilifolium. This individual is Tall thoroughwort (Eupatorium altissimum), a species with a strong preference for higher pH soils and dry, sunny conditions. E. perfoliatum and E. serotinum both prefer moister conditions, and the only other to also occur on dry, higher-pH sites, E. sessifolium, prefers greater shade. The combination of dry, high pH conditions and full sun makes this species the most likely in its genus. Knowing that a site has high soil pH can be as useful as morphology. But conversely, when you have definitively ID'ed a plant as this species, it can function as a clue to the soil conditions, which may help you to identify other plants on the same site. Photo © Tom Field, CC BY 4.0, Source.If using keys or references that do not mention habitat except in passing, I recommend supplementing them with other sources that discuss habitat more deeply, as part of the ID process. Often, considering habitat can confirm a correct ID, or can prompt you to question an incorrect ID, when you would have remained uncertain if considering morphology alone.
Beware planted or tended plants: habitat cannot be used to ID them!
A caveat to the advice to consider habitat is that habitat is only useful to identify wild plants. Plants that are cultivated, i.e. grown intentionally in gardens or landscaping, may grow in conditions that they would rarely or never survive in without help.Irrigation and/or mulching can allow moisture-demanding plants to survive on drier sites. Fertilization and/or mulching can allow plants to grow in soil lacking key nutrients. Wrapping and/or planting near a heated building can allow plants to overwinter in colder climates. Some plants can survive on a site even if they would never be able to germinate and establish there, and thus can be found if planted on sites where they would never establish on their own. And protecting plants from herbivory and/or pathogens through fencing and/or chemical treatments can allow stressed plants to survive in conditions they are not adapted to.
Perhaps most importantly of all, gardens remove competition. There are many examples of plants that prefer somehow adverse conditions and are not competitive in "ideal" conditions. Nearly all plants can survive in moist, well-drained conditions and slightly acidic pH (around 6.0.) However, plants adapted to drought, poor drainage, strongly acidic, or alkaline soil invest their energy into costly adaptations that allow them to survive these harsher conditions. In the wild, they would be overtaken by other plants that forgo these adaptations and instead grow faster in ideal conditions.
 This photo illustrates the perils of using habitat to identify garden plants. Here, a northern blue flag (Iris versicolor) is growing on dry ground on the north side of a house. This species normally only occurs in wetlands with standing water. The mulch retains water which allows it to survive on drier ground, and having it planted on the north side of the house further reduces drought stress. However, in the wild it usually is found on sites with much more light. But here, competing vegetation is also kept away from it, ensuring that it gets all of the relatively little sunlight shining on this area. Both factors likely allow it to survive on a drier, less-sunny site than it would normally occur on. Photo © Daveozric, CC BY 4.0, Source.
This photo illustrates the perils of using habitat to identify garden plants. Here, a northern blue flag (Iris versicolor) is growing on dry ground on the north side of a house. This species normally only occurs in wetlands with standing water. The mulch retains water which allows it to survive on drier ground, and having it planted on the north side of the house further reduces drought stress. However, in the wild it usually is found on sites with much more light. But here, competing vegetation is also kept away from it, ensuring that it gets all of the relatively little sunlight shining on this area. Both factors likely allow it to survive on a drier, less-sunny site than it would normally occur on. Photo © Daveozric, CC BY 4.0, Source.For these reasons, habitat cannot be used to identify planted and/or cultivated plants. However, the keyfactor is whether a plant is planted and/or tended, not where it is growing. Gardens and manicured landscapes are often full of wild plants, and these anthropogenic habitats can provide accurate habitat clues for identifying wild plants, a.k.a. "weeds", that come up in them. In fact, there are large numbers of plants in North America that primarily (and sometimes only) occur in anthropogenic habitats, and indeed, lawns and gardens are some of the most common habitats in which certain plants grow. And conversely, you can find planted plants in the wild, especially where a previously tended garden has become abandoned and overtaken with wild forest, leaving a few surviving trees, shrubs, or understory plants which may not be reproducing in the wild.
So, the key to knowing how to use habitat as an ID clue is to learn the distinction between a wild plant (which may or may not occur in a garden) and a planted or cultivated plant (which may or may not occur in a wild ecosystem.)
Once you've narrowed it to a reasonable number of species, read descriptions and look up pictures of those species.
When you've used the techniques above to narrow your search down to a manageable number of species, the next step is to study the potential species and see if any fit. One of the best resources for verbal descriptions is Flora of North America (FNA). Unfortunately, FNA is not complete, although over time it is becoming more so. FNA has detailed botanical descriptions of each species as well as higher taxa. It is particularly useful in the case where you want to check specific, observable traits, like counting or measuring or checking the texture of various parts of the plant. FNA uses a lot of specialized terminology so if you have not taken a course in or otherwise learned this terminology, you may need to look up a lot of terms. There are many local and regional flora, like the book Plants of Pennsylvania that have similar descriptions. That particular book also has an outstanding glossary, parts of which are illustrated, so if you live in or near PA I would recommend buying it.Looking up pictures of various species can be valuable, but also tricky. I recommend avoiding using a generic image search, as misidentifications are common on the internet, especially for tough-to-identify species. The best sources for images are dedicated plant websites where the maintainers have rigorously checked the ID of each photo before including it in their gallery, making misidentifications rarer. Start with a local source for your region, like Illinois Wildflowers, Maryland Biodiversity Project, or Calflora, or branch out to a regional one like FSUS.
User-uploaded sources such as iNaturalist can also be a source of images, but the potential for errors is higher on these sources. I recommend only using iNaturalist for images only if you are comfortable using descriptions in sources like FNA to double-check an ID.
Most people will want to look at pictures of live plants as they occur in the wild, but for certain specialized checks, you can also look up photos of herbarium specimens. Because herbarium specimens can be quite old, make sure to check for any taxonomic changes or alternate classification schemes. For example, many herbarium specimens of tall goldenrod (Solidago altissima) are listed under the name Solidago canadensis subsp. altissima.
Let your brain process the information.
Learning takes time and you can benefit from taking breaks and revisiting the material multiple times. Once you've checked descriptions and photos and given your brain a few days to process the information, go back outdoors and look at the mystery plant again. You may have to go between the plant and the online or text sources multiple times, each time finding some new trait to examine. Over time, you will not only get better at distinguishing plants, but you will begin to connect the specialized terminology with the up-close view of the plants. Over time, these connections will make keys and botanical descriptions more accessible.Check for common lookalikes.
If you think you may have identified something, but are not 100% sure, you can check for common lookalikes. Print books on plant identification often list lookalikes in the entry for each plant. Again, one of the best sources is iNaturalist. On iNaturalist, on the page for a particular species, there is a "Similar Species" tab. Furthermore, the listings on this tab can be filtered by region; in the top-right of the page, you can change the region and you will get region-specific listings.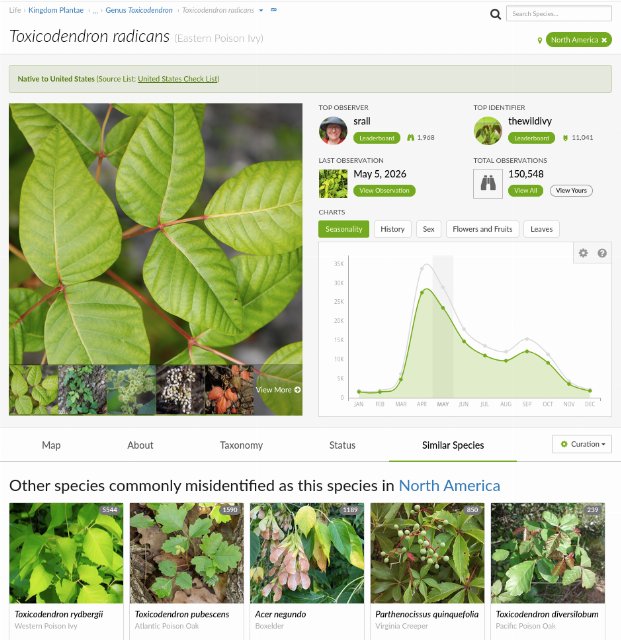 This screenshot of the iNaturalist page for Eastern Poison Ivy shows the "Similar Species" tab. In this case, the two most commonly-confused species are in the same genus (and pose a similar risk of causing a rash) but the third-most-confused is a species of maple that does not cause this rash, and the fourth is a vine that is also not closely related.
This screenshot of the iNaturalist page for Eastern Poison Ivy shows the "Similar Species" tab. In this case, the two most commonly-confused species are in the same genus (and pose a similar risk of causing a rash) but the third-most-confused is a species of maple that does not cause this rash, and the fourth is a vine that is also not closely related.Lookalikes are often, but not always closely-related. For example, for eastern poison ivy (Toxicodendron radicans), the most common lookalikes are western poison ivy (Toxicodendron rydbergii) and Atlantic poison oak (Toxicodendron pubescens), but the third-most-confused plant is box elder (Acer negundo), which is not closely related and is actually a type of maple (Acer sp.), and the fourth is Virginia creeper (Parthenocissus quinquefolia), which is more closely related to grapes than poison ivy. Checking these listings may give you ideas of lookalikes that you did not think to check. Our plant articles also contain a "Similar Plants" section but it is nowhere near as complete as iNaturalist's listings.
Regional web sources also frequently list common lookalikes. Illinois Wildflowers, Minnesota Wildflowers, and Go Botany (for New England) are three of the best regional resources that commonly list lookalikes and explain how to distinguish them.
To be truly rigorous, you will need to check options that do not appear in these lists, but the lists can be useful early in the process. You also can learn about plants you did not know about from these lists.
If you can't get started, AI can make initial guesses, but must be checked rigorously.
Sometimes you see a plant and you are just stumped. In these cases, AI-based apps can sometimes get you a starting point. Usually, people use these as smartphone apps, but some can also be accessed as websites. We recommend iNaturalist, Pl@ntNet, plant.id by Kindwise, and Flora Incognita for this purpose, for reasons we explain below. However, even the best apps make mistakes and are only suitable for initial guesses.AI is inherently unreliable and AI suggestions need to be checked rigorously.
At least in its current manifestations of LLM's, AI is notoriously unreliable, and has been known to hallucinate (outright make-up information) as well as to reflect existing biases. Furthermore, there is evidence that many AI models have been getting worse over time in part due to being trained on AI-generated content.And yes, there is AI-generated plant ecology content on the web. While researching plant distribution, ecology, and taxonomy, I have come across whole websites with databases of plants and articles about them, including complete fabrications, like saying particular species occur in certain locations where no reliable sources say they do. And even before the flood of AI slop to wade through, the web has long been filled with horticultural and gardening websites that take a casual approach to both plant identification and taxonomy, such as the fiasco of the invasive Asian jumpseed (Persicaria filiformis) being labeled and sold as jumpseed (Persicaria virginiana).
Any AI model is only as good as the dataset it is trained on, and this fact is as true for plant ID as any other task one might give it. There aren't many plant experts out there, and most of them aren't spending the bulk of their time training and refining AI models. As such, the apps used for plant ID make frequent mistakes, and the mistakes mimic the errors that humans frequently make, including misidentifications based on visually-similar plants, and confusion based on reclassifications or taxonomic inconsistencies. Poorly-designed AI's will fare even worse, reflecting misidentifications taken from unreliable websites and often just making bizarre guesses that have you scratching your head about how they might have occurred.
Which AI tools are best?
We recommend four AI-based tools for plant identification: Pl@ntNet, iNaturalist, plant.id by Kindwise, and Flora Incognita. All four are available as smartphone apps, and all but Flora Incognita also are usable as websites. iNaturalist requires registration but the others are usable as a "guest". A caveat is that I recommend the full iNaturalist app, not their simplified "Seek" app. The Seek app not only does not produce as accurate ID's, but it also lacks many of the interactive features that make iNaturalist most useful (even for ID.)Pl@ntNet was launched in 2009 and is run by a consortium of four research organizations and a foundation. iNaturalist was launched in 2008 and is run as a non-profit and is currently administered as a joint effort of the California Academy of Sciences and the National Geographic Society. Flora Incognita was launched in 2014 and is a joint project of the Technische Universität Ilmenau and the Max Planck Institute. All of these projects are open about their team of people and general methodology, contrasting with most other plant ID apps which share little information. Of the four, iNaturalist has the advantage of being integrated with community ID's and social features which not only facilitate conversations that can help you learn how to ID plants, but also lead the ID's (and AI) to become more accurate over time. Kindwise was recognized in a 2020 study as being the best of 10 free AI plant ID apps.
One of the best-known apps for plant ID is called Picture This. Picture This has a lot of rave reviews online, and claims to include "botanical experts" on their "team", but they do not even list a single person's name, nor do they explain anything about the source of their data used to train their models, let alone their methodology. I would put Picture This and any similar apps in the "use at your own risk" category.
Curious to see whether my judgment on these apps was accurate, I decided to give a test to a bunch of apps, including our four recommended ones and three others that I would not recommend. The test involved feeding them a picture of Asian jumpseed (Persicaria filiformis), which is a good indicator of whether a model has been trained based on rigorous identification using up-to-date taxonomy. A model trained on outdated sources, or on the web at large, would be likely to misidentify this plant as Persicaria virginiana because the web is dominated by nurseries and horticultural sites that label it as such.
 This naturalized population of Asian jumpseed (Persicaria filiformis) is growing in Mississauga, ON. Most of these populations originated as escapes of the 'Painter's Palette' cultivar which was mislabeled as jumpseed (Persicaria virginiana); many gardeners bought this cultivar and planted it thinking they were planting a native plant, only to contribute to introducing a new invasive to North America. Public Domain photo by Reuven Martin, Source.
This naturalized population of Asian jumpseed (Persicaria filiformis) is growing in Mississauga, ON. Most of these populations originated as escapes of the 'Painter's Palette' cultivar which was mislabeled as jumpseed (Persicaria virginiana); many gardeners bought this cultivar and planted it thinking they were planting a native plant, only to contribute to introducing a new invasive to North America. Public Domain photo by Reuven Martin, Source.Test Results
- iNaturalist - PASS - correctly guesses Persicaria filiformis as the first guess and presents Persicaria virginiana as the second guess.
- Pl@ntNet - PASS - correctly guesses Persicaria filiformis as the first guess, giving it an 83% probability and 16% to Persicaria virginiana. Showing the percentages is a nice touch, providing information that iNaturalist does not.
- Flora Incognita - PASS - correctly guesses Persicaria filiformis, also boldly giving a claim of 99% confidence.
- plant.id by Kindwise (formerly FlowerChecker) - PASS - Correctly guesses Persicaria filiformis.
- Picture This - FAIL - incorrectly guesses Jumpseed - Persicaria virginiana.
- Snap Plant - FAIL - guesses Persicaria maculosa, so it gets the genus correct but the species wrong.
- plantid.in - EPIC FAIL - gives the wildly-wrong guess of American burnweed (Erechtites hieraciifolius), a real head-scratcher as to how it came up with this.
This example used a crisp photo of a plant with distinctive foliage, and it is a species that not only is sold in nurseries but has gotten attention due to being an invasive that was mislabeled as a native, leading reputable sources to prioritize updating taxonomies and correcting misidentifications. Many species are both harder to ID and get a lot less attention than this one does. I frequently see iNaturalist make incorrect guesses as their first guess, and I have seen other online sources that have described all three of our other recommended apps also making incorrect guesses. So, even if you are working with the apps that seem to be the best options here, you still need to see them only as a starting point.
Keep in mind that the different apps can sometimes be better at different subsets of plants, so if you find one of them isn't getting you any closer to a correct ID, you could try another one from the subset of reputable apps. The goal is to get you a guess that is correct to at least genus or family level, when you have otherwise reached a dead-end in your ID attempts, so you might as well use one after another and see if any of them can lead you to a correct ID. But always go back to the reliable sources and check the ID thoroughly, even if it looks right at a glance.
Try it out!
Can you identify the plant at the start of this post? Or better yet, how about just go outside and try to identify anything you find? Or still better, find a plant you already recognize and observe it more closely, and then come back inside and look it up online and read more about it and research its relatives and lookalikes found in your area. Don't start with AI tools; start by building off what you know, researching more in reliable sources, and connecting that with real-world observation.The Bias Against Tall Plants
November 11th, 2025 by Alex Zorach
In our October 2024 post we noted that gardening and landscaping practices have ecological consequences, especially as an increasing portion of our land consists of managed landscapes. Gardening and landscaping practices are subject to cultural norms, which can include long-standing beliefs as well as fads and trends. These norms also influence the management of wild lands, especially at edges and other interfaces between wild and managed lands such as are common in parks.Currently, the concept of native plants is trending, in part due to the popularity of the books by Doug Tallamy. But even though the concept of native plants has broken into the mainstream, numerous ecological concepts are going under-appreciated, and numerous biases, many of which have negative ecological consequences, are persisting.
 Western society has glorified the aesthetics of gardens like this, with short plants and lots of space. Such gardens are practical only in more arid regions and on sites with poor soil. This garden is located in San Mateo, CA, in a region with a long dry season in summer. This aesthetic works better here than in the more humid portions of the country where the conditions support taller plants. Photo © Jeff Silva, California Native Plant Society, CC BY 2.0, Source.
Western society has glorified the aesthetics of gardens like this, with short plants and lots of space. Such gardens are practical only in more arid regions and on sites with poor soil. This garden is located in San Mateo, CA, in a region with a long dry season in summer. This aesthetic works better here than in the more humid portions of the country where the conditions support taller plants. Photo © Jeff Silva, California Native Plant Society, CC BY 2.0, Source.This post tackles a particular bias: the bias against "tall" plants, and its counterpart, the bias in favor of "short" plants. This bias goes hand-in-hand with lawn culture, and has consequences diverse areas, including:
- proliferation of invasive plants
- weakening of native plants by breeding cultivars to be shorter
- failure of ecological restoration projects because of selecting shorter plants on sites suited to taller plants
- degradation of edge habitats by selective removal of herbaceous plants that are "too tall"
The Ecology of Plant Height
Plants' physical attributes, including height, are a function of their ecology, reflecting adaptation to their habitats. Habitat includes both the environmental conditions (such as moisture and light availability, and soil texture and chemistry) and other organisms present in the ecosystem, including both animals and competing plants.In order to grow taller, herbaceous plants need resources, including water, nutrients, and light. However, growing taller is still costly, so it is only advantageous if it offers something that a short habit does not. The main advantage of height is pushing up through competing vegetation to reach more light. Competition not only increases the reward of height, but also facilitates it, since plants can lean on each other for structure. Many tall plants cannot support their own weight alone, staying upright and pushing up through other vegetation only by leaning against each other. As such, most tall plants occur in communities of other tall plants.
 This photo shows a tallgrass prairie at Midewin National Tallgrass Prairie in Illinois. A tallgrass prairie is a plant community of tall plants; many of these plants cannot support their own weight if growing in isolation, but in a dense community like this they can lean on each other for support. Photo © US Forest Service, Public Domain, Source.
This photo shows a tallgrass prairie at Midewin National Tallgrass Prairie in Illinois. A tallgrass prairie is a plant community of tall plants; many of these plants cannot support their own weight if growing in isolation, but in a dense community like this they can lean on each other for support. Photo © US Forest Service, Public Domain, Source.Besides the increased light capture, there are secondary benefits of height, including visibility of flowers to pollinators, and in wind-pollinated plants, increased reach of pollen and ability to capture pollen from distant plants. Wind-dispersed seeds can also spread over longer distances. These wind-facilitated benefits though come with a price: the higher wind speeds at greater heights worsens drought stress, thus increasing water demands more than would be expected by light exposure and leaf surface area alone.
 This inflorescence on Canada goldenrod (Solidago canadensis) is covered with clusters of mature seeds, each attached to a pappus, a structure designed to carry the seed away in the wind. As wind speeds increase with height, the height of this plant is directly related to the distances its seeds are able to disperse over. Photo © Robb Hannawacker, Public Domain, Source.
This inflorescence on Canada goldenrod (Solidago canadensis) is covered with clusters of mature seeds, each attached to a pappus, a structure designed to carry the seed away in the wind. As wind speeds increase with height, the height of this plant is directly related to the distances its seeds are able to disperse over. Photo © Robb Hannawacker, Public Domain, Source.For these reasons, most tall herbaceous plants occur in habitats where there is abundant water, light, and nutrients, and heavy ground-level competition. These habitats include tallgrass prairie, wet meadows and openings in floodplain forests, rich, moist forest edge habitats (including streambanks and shores of lakes, ponds, and open wetlands, as well as anthropogenic forest edges), swamps, and locally-disturbed sites in bottomland forests or rich mesic upland forests. In drier habitats and on sites with poorer soil, the drought stress, lack of competition, and scarcity of nutrients combine to favor shorter plants. On shadier sites such as a closed-canopy forest, low-light conditions favor either woody plants like trees or shrubs, or low-growing herbaceous plants.
Much of North America's population is found in areas favoring tall herbaceous plants.
The human population in North America concentrates along the humid coasts where forests were dominant, and into the eastern portion of the Great Plains, where tallgrass prairies were dominant. Furthermore, in forested regions, humans tend to settle near sources of water and in areas with richer soil, as these areas support agriculture capable of feeding larger populations.Humans often increase the habitat for tall herbaceous plants.
Without the influence of humans, much of the Eastern Temperate Forests would support closed-canopy forests in which there are fewer tall herbaceous plants. In many of these regions, historically, Native Americans practiced controlled burns which favored savannas, which supported more tall herbaceous plants, including both grasses and broadleaf plants. European colonization brought the clearing of forests and conversion of land to agriculture, and tall plants proliferated on cropland margins, fallow fields, and in the early-successional growth on sites where agriculture was abandoned and forest was allowed to grow up again. In the present day, an abundance of roads and paths and the fragmenting of forests has created more edge habitat, where higher levels of light reach the ground underneath trees, and tall herbaceous plants thrive.Resisting a natural equilibrium has a cost.
I hope I've convinced you that, at least some of the time and on some sites, the conditions favor tall plants. Attempting to resist such a natural equilibrium comes with its costs, which the rest of this post will explore.Leaving The Tall Plant Niches Vulnerable to Invasion
If you remove a certain species from an ecosystem, its niche will usually be colonized by some other plant. Removing a tall native plant will usually lead its niche to be colonized by a plant of similar height. However, if over time, tall native plants are repeatedly and systematically removed from ecosystems, their niches will be colonized either by shorter native plants, or by non-native plants. Because tall plants tend to be more competitive than short plants on sites rich enough to support tall plants, systematic removal of tall native plants can lead a site to be more vulnerable to invasive plants.Many of the invasive plants that fill these niches are themselves tall. But the bias against tall plants acts on them as well. People are more likely to notice tall invasives and target them for removal, and much of the management of land is not done with respect to a plant's native status, so the bias against tall plants acts equally on tall invasives. When all tall plants are excluded, they become replaced by shorter vegetation. And as I explain below, there are some systemic reasons why this shorter vegetation is more likely to be dominated by introduced plants.
Proliferation of Invasive Plants Selected for Shortness
One pattern I have noticed about invasive plants in North America is that a surprisingly large portion of the worst invasive species have a low-growing habit. This pattern is the opposite of what one would expect based on competition, as tall plants often out-compete shorter plants. Although there are more examples of such low-growing invasive plants in the humid east, this problem is not restricted to that region. The southwest has crystalline ice plant (Mesembryanthemum crystallinum) and slenderleaf iceplant (Mesembryanthemum nodiflorum), and fire-prone areas of the interior west and western great plains have cheatgrass (Bromus tectorum), both of which are lower-growing than many of the plants in the ecosystems they tend to invade.The low height of these invasive plants tends to create other ecological problems, beyond the typical collapse of the food web caused by non-native species being eaten by fewer insect specialists. The food web problem is not directly related to height. However, another serious problem, erosion, is directly related. Although there are some exceptions, on average, shorter or smaller plants tend to have shallower and less-extensive root systems than tall plants. And, as-such, the proliferation of short or low-growing invasive plants tends to worsen erosion and increase the risk of larger-scale mass wasting such as slumping of soil.
 Crystalline ice plant (Mesembryanthemum crystallinum) is an invasive plant with a low-growing habit that outcompetes even taller vegetation in Mediterranean California through mechanisms such as inhibiting seed germination and increasing the soil's surface salinity. It is shallow-rooted, so its dominance leads to dead zones with large areas of exposed soil, creating an erosion problem on slopes like pictured here. The native vegetation on slopes like this would be dense chaparral several feet in height, with deep roots that stabilize the soil. Photo © mel letterman, CC BY 4.0, Source.
Crystalline ice plant (Mesembryanthemum crystallinum) is an invasive plant with a low-growing habit that outcompetes even taller vegetation in Mediterranean California through mechanisms such as inhibiting seed germination and increasing the soil's surface salinity. It is shallow-rooted, so its dominance leads to dead zones with large areas of exposed soil, creating an erosion problem on slopes like pictured here. The native vegetation on slopes like this would be dense chaparral several feet in height, with deep roots that stabilize the soil. Photo © mel letterman, CC BY 4.0, Source.Shorter plants also tend to have less biomass and thus, when they dominate ecosystems and out-compete taller plants, they reduce the total extent of the ecosystem, which reduces not only food available to animals, but also other ecosystem services plants provide, including cover for animals, solar capture, reduction of wind speed, and evapotranspiration, which in turn provides both temperature regulation and runoff reduction. The replacement of tall native vegetation by short invasives causes a cascade of harm extending beyond the ecosystem itself, leading to a reduction in climate regulation in the surroundings, harm to wildlife that spends only a portion of time in the ecosystem, and harm to waterways downstream.
The reason there are so many low-growing invasive plants is that humans have imported these plants to the continent disproportionately. One of the main ways short plants get introduced is through lawns, so our first examples of invasive plants will be forms of turfgrass.
Turfgrass
A large portion of the introduced grasses in North America were brought here as turfgrass. Although states and other governments are reluctant to label these plants as invasive, the reasons for doing so are more economic and cultural than ecological. One of the most widely-planted turfgrasses in the US is tall fescue (Lolium arundinaceum), which also happens to be one of the most aggressive in the wild. The USDA's FEIS acknowledges "Tall fescue can be invasive in native vegetation," giving the example of the Clymer Meadow Preserve in Texas, where it is encroaching on native vegetation, and notes "Tall fescue has devastated many other prairie remnants in Texas and to the north." This tall fescue (Lolium arundinaceum) is forming a monoculture at Bozeman Sculpture Park in Bozeman, MT. These monocultures shut out nearly all native vegetation. This species primarily spreads through lawns and pastures, and is often still planted intentionally. Photo © Matt Lavin, CC BY 4.0, Source.
This tall fescue (Lolium arundinaceum) is forming a monoculture at Bozeman Sculpture Park in Bozeman, MT. These monocultures shut out nearly all native vegetation. This species primarily spreads through lawns and pastures, and is often still planted intentionally. Photo © Matt Lavin, CC BY 4.0, Source.Some other turfgrasses that can become invasive include bermudagrass (Cynodon dactylon), Kentucky bluegrass (Poa pratensis) (which, contrary to its name, is not from Kentucky), rough bluegrass (Poa trivialis), perennial ryegrass (Lolium perenne), italian rye-grass (Lolium multiflorum), hard fescue (Festuca trachyphylla), creeping bentgrass (Agrostis stolonifera), bahiagrass (Paspalum notatum), centipede grass (Eremochloa ophiuroides), and Korean lawngrass (Zoysia japonica). This list is hardly exhaustive.
So many turfgrasses are invasive because they been bred over many generations both to be vigorous in North America's climates and soils, to be competitive against "weeds" (i.e. any other plant that might compete with them), and to resist any insect "pests" that might eat them. All three factors increase the likelihood of a species becoming invasive.
Groundcovers
Groundcovers are low-growing plants, typically either vines or herbaceous plants that reproduce by rhizomes or stolons, and that stay relatively low to the ground without needing to be mowed. Some grow taller than a typical lawn, but most are shorter than about 18 inches (1.5 feet) and some are much shorter. Many of them are evergreen. Some are succulents. Some have showy flowers. It is important to understand that groundcover is a horticultural term, not an ecological one: it refers to a plant's use in the garden, not its natural growth habit in the wild.Some of the worst invasive plants in North America were introduced by being widely planted as groundcovers. Such examples include English ivy (Hedera helix), lesser celandine (Ranunculus ficaria), common iceplant (Mesembryanthemum crystallinum), winter creeper (Euonymus fortunei), Japanese pachysandra (Pachysandra terminalis), creeping jenny (Lysimachia nummularia), common periwinkle (Vinca minor), bigleaf periwinkle (Vinca major), stringy stonecrop (Sedum sarmentosum), and goldmoss stonecrop (Sedum acre).
 This open woodland in Decatur, Georgia, is completely overrun with english ivy (Hedera helix), which not only forms a monoculture over the entire ground, but also covers most of the tree trunks. English ivy is particularly pernicious as its seeds are bird-distributed and its seedlings establish readily in shade, so it is not limited to vegetative spread. Most infestations originate in landscaping where it is widely used as a groundcover; note the homes in the background. Ivy is often most dominant in small woodland fragments in urban and suburban areas. Photo © Emily Summerbell, CC BY 4.0, Source.
This open woodland in Decatur, Georgia, is completely overrun with english ivy (Hedera helix), which not only forms a monoculture over the entire ground, but also covers most of the tree trunks. English ivy is particularly pernicious as its seeds are bird-distributed and its seedlings establish readily in shade, so it is not limited to vegetative spread. Most infestations originate in landscaping where it is widely used as a groundcover; note the homes in the background. Ivy is often most dominant in small woodland fragments in urban and suburban areas. Photo © Emily Summerbell, CC BY 4.0, Source.Other groundcovers that have established in the wild but are not quite as aggressive include spotted henbit (Lamium maculatum), green hellebore (Helleborus viridis), bugleweed (Ajuga reptans), common houseleek (Sempervivum tectorum), evergreen candytuft (Iberis sempervirens), and several more stonecrops including orpin aizoon (Phedimus aizoon), white stonecrop (Sedum album), spanish stonecrop (Sedum hispanicum), European stonecrop (Sedum ochroleucum), and tasteless stonecrop (Sedum sexangulare). These plants can still be aggressive in some contexts, and any of them has the potential risk of becoming invasive at some point in the future.
There are likely many other plants with potential to become invasive if widely planted as groundcovers, but that have not yet become so simply because they have not been widely planted and/or bred. An example of a plant that is currently in the process of becoming more invasive is creeping liriope (Liriope spicata), which was widely planted as a replacement for English ivy, a decision that was both short-sighted and lacking in ecological awareness.
As with turfgrasses, groundcovers are also bred for traits that increase their likelihood of becoming invasive: tolerance of North America's climate and soil conditions, resistance to invasion by "weeds", and insect resistance.
Weedy low-growing plants that survive in lawns
Not all the introduced low-growing plants ended up here intentionally. A large portion of them propagate and spread as lawn weeds. Lawns provide an ideal habitat for the spread of such plants, not only because the mowing removes any taller competition, but also because the lawn equipment itself spreads plant seeds around, both locally and to new sites when the seeds ride on the equipment.Introduced weedy plants that grow in lawns include unwanted grasses, such as annual bluegrass (Poa annua), hairy crabgrass (Digitaria sanguinalis), smooth crabgrass (Digitaria ischaemum), quackgrass (Elymus repens), awnless brome (Bromus inermis), plantain signalgrass (Brachiaria plantaginea), and indian goosegrass (Eleusine indica). Some of the turfgrasses discussed above, which may be intentionally planted on some sites, can also be unwanted weeds in other lawns. Such sometimes-unwanted grasses include tall fescue, creeping bentgrass, and rough bluegrass.
The list of introduced broadleaf weedy plants that thrive in lawns is even longer. Some of the most common ones include lesser celandine, mock strawberry (Duchesnea indica), European cinquefoil (Potentilla reptans), birdeye speedwell (Veronica persica), common plantain (Plantago major), narrowleaf plantain (Plantago lanceolata), common dandelion (Taraxacum officinale), scarlet pimpernel (Lysimachia arvensis), white clover (Trifolium repens), japanese clover (Kummerowia striata), korean clover (Kummerowia stipulacea), suckling clover (Trifolium dubium), black medick (Medicago lupulina), bird's-foot trefoil (Lotus corniculatus), ground ivy (Glechoma hederacea), common chickweed (Stellaria media), henbit deadnettle (Lamium amplexicaule), purple deadnettle (Lamium purpureum), and low smartweed (Persicaria longiseta). Again, this list is far from comprehensive; a full list for North America would be pages long.
 Korean clover (Kummerowia stipulacea) is one of many introduced plants that spreads in part as a lawn weed, but also colonizes other habitats where it takes the place of native vegetation. Here it is growing in exposed gravel in Capon Bridge, WV, along the Cacapon river. Note that nothing is growing in with it; although it is a nitrogen fixer, it tends to form monocultures. Photo © Matthew Gerke, CC BY 4.0, Source.
Korean clover (Kummerowia stipulacea) is one of many introduced plants that spreads in part as a lawn weed, but also colonizes other habitats where it takes the place of native vegetation. Here it is growing in exposed gravel in Capon Bridge, WV, along the Cacapon river. Note that nothing is growing in with it; although it is a nitrogen fixer, it tends to form monocultures. Photo © Matthew Gerke, CC BY 4.0, Source.Although people are not actively breeding these weeds to be more invasive, lawn culture unintentionally has this effect. In nature, many of the habitats that these plants would colonize are ephemeral, persisting only temporarily in response to a disturbance. But the regular mowing of lawns creates a new, artificial habitat in which these plants can produce generation after generation, persisting in this habitat consistently and long-term, often in large numbers.
So, instead of having a small number of plants producing seed and then dying out after a year or two, only to have suitable habitat crop up, potentially farther away, potentially years later, we now have a scenario in which the plants are breeding in large numbers every year. The massive amount of lawn further increases the population of these plants, and the fact that lawn equipment can move seeds from site to site, increases the speed with which genetic material spreads over large regions. All of these factors make the populations more vigorous and allow them to adapt to the conditions on this continent, increasing the likelihood that these plants become invasive.
Weakening of native plants by breeding short cultivars
Modern horticulture, landscaping, and gardening practices don't just introduce new, potentially invasive plants, but also alter our native plants. The horticulture industry breeds cultivars to have "desirable" traits in gardens. Modified traits may include flower color and structure, foliage color, and growth habit. One of the most common modifications made with such breeding is to decrease the height of plants.Above we explained how a plant's height is a key aspect of its fitness and adaptation to its habitat. Modifying a plant's height by selective breeding makes it less fit in the habitats to which it is otherwise adapted. Shortened plants tend to be less competitive with other vegetation, and are usually less shade tolerant as the upper leaves tend to capture the most light. Shortened plants may also be less able to quickly and successfully cross-pollinate, whether the plant is wind- or insect-pollinated, and less able to distribute its seed larger distances particularly if its seed is wind-distributed. Although there are also potential benefits of shorter height (such as greater tolerance of drought and low-nutrient conditions), altering a plant from its equilibrium under natural selection tends to decrease its fitness. If it were beneficial to be shorter, it would have already become shorter through natural selection.
Plants also have some morphological plasticity, meaning that, even plants of identical genetics adjust their characteristics (including height) dynamically based on conditions. Cultivars are not just bred for specific traits, they are also typically bred to have less variation in those traits on different sites, so they are more "predictable" in the garden. In other words, their morphological plasticity is reduced, which also reduces their fitness in the wild.
One such cultivar is 'Little Joe' cultivar of coastal plain joe pye weed (Eutrochium dubium). Although E. dubium is naturally the shortest of the five Eutrochium species, this cultivar has been bred to make its maximum height even shorter: it grows only to about 4 feet, contrasting with the wild forms of the plant, which commonly grow to about 5 feet. This cultivar is also a classic example of the decrease in variability: both the cultivar and wild forms range in height, and can be as short as 3 feet at maturity. But the cultivar ranges from 3-4 feet in height, whereas the wild form ranges from 3-5 feet. The difference of that missing last foot of height tends to occur on plants grown in the conditions with greater moisture and nutrients, conditions where the plant is most likely to face competition from other plants.
 Although coastal plain joe pye weed (Eutrochium dubium) is naturally the shortest of the Eutrochium species, its wild-type plants still grow up to 5 feet tall in typical conditions; this height is necessary for them to compete with the other vegetation in the wetlands they inhabit. Photo © Bonnie Semmling, CC BY 4.0, Source.
Although coastal plain joe pye weed (Eutrochium dubium) is naturally the shortest of the Eutrochium species, its wild-type plants still grow up to 5 feet tall in typical conditions; this height is necessary for them to compete with the other vegetation in the wetlands they inhabit. Photo © Bonnie Semmling, CC BY 4.0, Source.The genetics encoding for height of these cultivars get passed on to future generations. Although the plants do tend to revert back to a more wild-type plant eventually, the shortened height does tends to persist, somewhat unpredictably, through at least one or two generations. I have done a lot of ecological restoration work in New Castle County, Delaware, a region where E. dubium is both common in the wild, and where the 'Little Joe' cultivar is widely planted in gardens. E. dubium is a particularly good example of the effect of these cultivars on wild plants, because the species self-seeds prolifically both in wild ecosystems and in suburban gardens and weedy marginal areas in suburbs. And I have noticed a clear pattern of volunteer E. dubium near source populations of these short cultivars, themselves being shorter at maturity than volunteer plants of the same species found in more intact wild areas, such as along the lower Christina river where it is separated from a large chunk of forest in Lewden Greene Park, or adjacent to larger wetlands in Phillips Park, Newark.
Locally-sourced wild plants grown in gardens can protect the populations of those plants in nearby wild ecosystems, by seeding out into wild areas. The offspring of these garden plants compete more favorably against invasive plants in the wild because the wild populations have genetics that were the product of generations of natural selection for fitness in the wild. Such garden plants also serve as a reservoir for the genetics of the local wild populations, and can help preserve these genetics if a local wild population is temporarily extirpated. The importance of these reservoirs cannot be over-emphasized in a world where we have reduced wild land to a tiny proportion of its original extent, wild habitats are highly fragmented, and the local genetics of wild plant populations are at risk of being lost entirely.
When gardeners instead plant horticultural cultivars, it is not clear what the long-term effects on the local populations of these species will be. I have seen a lot of evidence to suggest that concern is warranted, and that these cultivars often produce seedlings that are less fit for survival in the wild. And excepting in the rare cases where cultivars are planted in the region from which their ancestors were sourced, they do not preserve the local genetics. There is also the possibility that a "worst case scenario" is also happening, in which the weakened cultivars are interbreeding with wild populations in ways that reduces their fitness, leading to decline of the populations of certain species, at least in some areas.
I have not yet seen any specific evidence that this is happening, but I also have not seen conclusive evidence that it is not. Concerningly, the horticulture industry is not studying any of these potential effects; they continue to breed and market and distribute plants primarily for their performance in gardens. What little consideration of ecology is being done has focused on the value of plants to insect populations (such as is studied at Mt. Cuba Center's trial garden.) But I have yet to see any research at all on the effect of cultivars on the genetics or fitness of local wild populations of plants.
Removal of Native Plants that Are "Too Tall"
The bias against tall plants affects property management both in home gardens and semi-wild spaces in managed parkland.A lot of people grow "native plant gardens" but manage them intensely, not allowing plants to reproduce freely. People who want to grow only shorter plants end up only growing a subset of plants adapted to drier, more nutrient-poor conditions. People sometimes remove even showy, visually-appealing plants such as cutleaf coneflower (Rudbeckia laciniata), tall goldenrod (Solidago altissima), and late boneset (Eupatorium serotinum), specifically because of their height.
 Tall plants like this late boneset (Eupatorium serotinum) are often singled out for control or excluded from gardens because of their height and tendency to flop into paths, as this particularly robust individual is doing here in Rochester, NY. Photo © Sandy Wolkenberg, CC BY 4.0, Source.
Tall plants like this late boneset (Eupatorium serotinum) are often singled out for control or excluded from gardens because of their height and tendency to flop into paths, as this particularly robust individual is doing here in Rochester, NY. Photo © Sandy Wolkenberg, CC BY 4.0, Source.The absence of tall native plants in suburban gardens is directly related to the ways in which rich, moist habitats, the ones where tall plants tend to naturally be dominant, are so overrun with invasives. An example of such habitats are wet meadows in floodplains, which are often common in suburban areas in land too low and flood-prone to build on. Many of these meadows are overrun with invasive plants, some of which colonize from seeds washed downstream from the suburban yards themselves. If people were less averse to planting tall plants, these meadows might have more intact ecosystems, populated by plants like cutleaf coneflower, wingstem (Verbesina alternifolia), and giant goldenrod (Solidago gigantea). The reduced height of vegetation from removing tall plants can create additional problems, increasing water speed during floods, which can increase flood severity downstream, and also increase erosion which causes soil loss upstream, and nutrient pollution, resulting in eutrophication, downstream.
Bias against tall plants in managed parkland can lead park staff to expend resources removing or killing native plants, driven either by complaints from the public, or just the discretion of the staff themselves. For example, in Newark, Delaware, I have seen park workers spray and kill American pokeweed (Phytolacca americana), Canada lettuce (Lactuca canadensis), and tall blue lettuce (Lactuca biennis), but leaving in place many shorter plants which were non-native such as prickly lettuce (Lactuca serriola) and the various sow thistles (Sonchus sp.) So the management was unintentionally favoring introduced vegetation. Changing these practices, however, is relatively easy and can have benefits.
 Wingstem (Verbesina alternifolia), a tall herbaceous plant, grows next to a managed edge between a parking lot and forest in Shenandoah National Park. Tall native plants along the edge reduce the potential for invasive plants and protect soil from erosion. The tall vegetation grows at a lower level where plants are less likely to flop onto the pavement, and this area is wetter from water draining off the road. This setup illustrates many aspects of good design and management, addressing multiple environmental and practical concerns together. Photo © Julia Beach, CC BY 4.0, Source.
Wingstem (Verbesina alternifolia), a tall herbaceous plant, grows next to a managed edge between a parking lot and forest in Shenandoah National Park. Tall native plants along the edge reduce the potential for invasive plants and protect soil from erosion. The tall vegetation grows at a lower level where plants are less likely to flop onto the pavement, and this area is wetter from water draining off the road. This setup illustrates many aspects of good design and management, addressing multiple environmental and practical concerns together. Photo © Julia Beach, CC BY 4.0, Source.During one year I went out with park staff along the James Hall Trail in Newark, DE and showed them how to distinguish the native lettuces from the invasive prickly lettuce. That year, the staff sprayed only the prickly lettuce, and a few of the native lettuces growing too close to the path, but left native lettuces that were not posing any risk of flopping into the path. In the next year, I saw a dramatic shift in the relative abundances of lettuce species along the trail, with the two native species becoming dominant, whereas in all prior years, the invasive prickly lettuce had been dominant. This change then produced other benefits. The native lettuces supported more aphids, which in turn supported other animals higher up on the food chain such as mantises and migrating warblers.
Failure of Ecological Restoration Projects
Projects can fail due to selection of short plants on sites better-suited to tall plants.
When I first started working in the area of ecological restoration, I was not prepared for just how much plant selection would be influenced by horticultural thinking and aesthetics. Pure ecological projects are rare. A large portion of ecological restoration projects are not conducted on truly wild land, they are only semi-wild landscapes in parks, historic preserves of old estates, or on grounds owned by schools, universities, cemeteries, or other organizations. They don't necessarily want a truly wild meadow, they want something that looks a certain way, and this "look" is plagued by the bias against tall plants. Johnsongrass (Sorghum halepense) is invading this meadow planting in Hillside Park in Newark, DE. This meadow, in a bottomland habitat around a pond, was planted with species adapted to a drier climate and conditions, such as hoary vervain (Verbena stricta) and lanceleaf tickseed (Coreopsis lanceolata). Species like this are often chosen due to their showy flowers and shorter size, even if they are not ideally suited to the site. Predictably, the site was invaded by taller plants. Had the meadow been planted with taller species native to bottomlands in this region, the johnsongrass might not have been able to invade. Note that the native tall goldenrod (Solidago altissima) has also colonized this site. Photo © Alex Zorach, CC BY 4.0, Source.
Johnsongrass (Sorghum halepense) is invading this meadow planting in Hillside Park in Newark, DE. This meadow, in a bottomland habitat around a pond, was planted with species adapted to a drier climate and conditions, such as hoary vervain (Verbena stricta) and lanceleaf tickseed (Coreopsis lanceolata). Species like this are often chosen due to their showy flowers and shorter size, even if they are not ideally suited to the site. Predictably, the site was invaded by taller plants. Had the meadow been planted with taller species native to bottomlands in this region, the johnsongrass might not have been able to invade. Note that the native tall goldenrod (Solidago altissima) has also colonized this site. Photo © Alex Zorach, CC BY 4.0, Source.Many times I have consulted with organizations looking to restore a site, come up with a plant list, and have some of the plants I or others recommended as "ecological powerhouses" be removed in the final list, often replaced by plants that are shorter and less vigorous. A common substitution is that people want the lower-growing butterfly milkweed (Asclepias tuberosa) in place of the taller, more aggressive common milkweed (Asclepias syriaca). Similarly, people often want to substitute shorter goldenrods such as gray goldenrod (Solidago nemoralis) or early goldenrod (Solidago juncea) for tall goldenrod (Solidago altissima), canada goldenrod (Solidago canadensis), or giant goldenrod (Solidago gigantea). Although these choices might work in a managed garden, they do not work in even a semi-wild landscape where plants freely seed in from the wind or bird droppings, and "duke it out", with the winners chosen by survival of the fittest.
Shorter plants only win the battle on sites with adverse conditions, like drought-prone coarse sandy soils, thin soil over a rock outcropping, or exposed mineral soil that is devoid of organic matter. But as I observed above, humans tend to live mostly on or near rich sites where the ecology favors tall plants, so restoration projects where the short plants are favored by conditions are the exception rather than the norm.
Predictably, the restored site becomes invaded by unwanted tall plants, often rhizomatous invasives such as common mugwort (Artemisia vulgaris), creeping thistle (Cirsium arvense), johnsongrass (Sorghum halepense), Japanese knotweed (Reynoutria japonica), or in the south, giant reed (Arundo donax). Sometimes unwanted native plants move in, such as common ragweed (Ambrosia artemisiifolia) or giant ragweed (Ambrosia trifida), both major allergens.
Degradation of Edge Habitats
The management of edge habitats poses an even more widespread problem than the half-hearted ecological restoration projects discussed above. Edge habitats include forests adjacent to parks or lawns on residential, commercial, industrial, or other institutional properties, as well as edges of roads and bike/pedestrian pathways that cut through forests. Many edge habitats are managed without any consideration of ecology whatsoever, and when it is considered at all, it is usually little more than an afterthought. Once again, the bias against tall plants rears its ugly head.Edge habitats are often managed by workers whose instructions or intentions are little more than to keep an edge "maintained" or "neat". As such, they have a great deal of discretion. They may also respond to complaints from the general public (especially in municipal parks) or the whims of management.
 These tall blue lettuce (Lactuca biennis) plants are growing along a roadside in Morse Township, MN, at an edge between the forest and the managed open habitat along the road. Tall plants like this naturally grow along many edge habitats, and help to protect the interior from wind as well as protecting the soil they grow in from erosion. Photo © Norma Malinowski, CC BY 4.0, Source.
These tall blue lettuce (Lactuca biennis) plants are growing along a roadside in Morse Township, MN, at an edge between the forest and the managed open habitat along the road. Tall plants like this naturally grow along many edge habitats, and help to protect the interior from wind as well as protecting the soil they grow in from erosion. Photo © Norma Malinowski, CC BY 4.0, Source.Tall plants also pose a practical management concern, in that they tend to flop into paths or roads. Forward-looking trail managers might want to act proactively, removing tall plants early, before they flop into a path. This removal can happen by mowing or bushwhacking farther back from the path than the standard mow line, by spot-treating plants behind the mow line with herbicide, or by weed-whacking or cutting isolated plants. Plants I have seen targeted in this way include lettuces, especially the native canada lettuce (Lactuca canadensis) and tall blue lettuce (Lactuca biennis) (which tend to grow taller than the invasive prickly lettuce (Lactuca serriola)), American pokeweed (Phytolacca americana), the taller species of goldenrod, common milkweed, and American burnweed (Erechtites hieraciifolius) (which is often mistaken for a thistle). Non-target plants are often killed in these control efforts, creating localized dead zones.
When tall plants are selected against in management of edge habitats, it can cause cascading problems in the interior of the habitat. Relative to open habitats, forest interiors have lower light levels, greater leaf litter, and a more moderate microclimate, with lower wind speeds and more regular temperatures. A natural forest edge produces a buffer of vegetation blocking both light and wind. This buffer consists both of vegetation from woody plants, such as trees and shrubs on the edge, and ground-level herbaceous plants. Edges tend to favor taller vegetation than forest interiors because of the higher light availability, and there is a bit of a positive feedback loop because taller ground-level vegetation captures more leaf litter around the base of plants, leading to high nutrient availability.
Removing the tall herbaceous plants right at the edge causes changes to the interior, farther back than the actively managed zone. Wind speeds increase, light increases, and temperature swings become more extreme. The removal of vegetation and increase in wind speed can cause leaf litter near the edge to blow away, exposing soil to erosion. These changes degrade the habitat for native plants and animals alike. Invasive shrubs and woody vines can colonize the zone set back from the edge in response to the higher light levels and disturbed soil. In the mid-Atlantic, such shrubs include Amur honeysuckle (Lonicera maackii) and burning bush (Euonymus alatus), and vines include Japanese honeysuckle (Lonicera japonica) and oriental bittersweet (Celastrus orbiculatus). The vines, and shrubs like Amur honeysuckle with a wide-arching habit, can root in a pocket of fertile soil and then extend their photosynthetic branches over areas with degraded soil. Once they establish in place of the native vegetation, they then begin the process of capturing leaf litter and nutrients, but creating a new structure that remains stable in the face of management practices, because they are rooted farther back, outside the zone of active management.
 This Japanese honeysuckle (Lonicera japonica) is growing on a roadside in Sherrill Township, Missouri, partly supported by a barbed-wire fence. Japanese honeysuckle, together with other invasive vines, often thrives along roadsides and other edge habitats where management practices remove ground-level herbaceous plants, but a vine rooted in a protected area farther back than the management zone is able to climb structures such as fences, trees, and shrubs, and then overhang the edge. Photo © Conway Hawn, CC BY 4.0, Source.
This Japanese honeysuckle (Lonicera japonica) is growing on a roadside in Sherrill Township, Missouri, partly supported by a barbed-wire fence. Japanese honeysuckle, together with other invasive vines, often thrives along roadsides and other edge habitats where management practices remove ground-level herbaceous plants, but a vine rooted in a protected area farther back than the management zone is able to climb structures such as fences, trees, and shrubs, and then overhang the edge. Photo © Conway Hawn, CC BY 4.0, Source.Without management that removes tall plants, there would still be invasive plants but they would find it more difficult to penetrate as far into the forest interior. Tall herbaceous plants tend to outcompete vines like Japanese honeysuckle, because the plant cannot reach a high permanent height without climbing woody vegetation, and herbaceous plants grow rapidly and shade out the honeysuckle's leaves. Removing the tall plants tends to make the honeysuckle more vigorous, as more light reaches the carpet of honeysuckle leaves growing on the ground.
Another problem with such removal is the creation of temporary dead zones, small patches which disturbance-loving plants can colonize. Depending on the timing of the control, there are a number of opportunistic invasive plants that can take advantage of these patches. Japanese stiltgrass (Microstegium vimineum) tends to colonize areas disturbed in spring to early summer, and short-lived plants like common groundsel (Senecio vulgaris) can colonize at just about any time of year. Edge habitats, which in our society are so prevalent that they form an interconnected network extending throughout whole regions, become corridors along which these invasive plants can spread.
 This Japanese stiltgrass (Microstegium vimineum) is growing along a roadside in Bluemont, VA. Stiltgrass often spreads along roads and paths, taking advantage of exposed soil from mowing, weedwhacking, or herbicide use. Stiltgrass will usually be excluded by tall perennial vegetation, as it is an annual with tiny seeds, and its seedlings cannot establish among heavy competition. Photo © Brian Magurn, CC BY 4.0, Source.
This Japanese stiltgrass (Microstegium vimineum) is growing along a roadside in Bluemont, VA. Stiltgrass often spreads along roads and paths, taking advantage of exposed soil from mowing, weedwhacking, or herbicide use. Stiltgrass will usually be excluded by tall perennial vegetation, as it is an annual with tiny seeds, and its seedlings cannot establish among heavy competition. Photo © Brian Magurn, CC BY 4.0, Source.The degradation of managed edge habitats, because it is such a widespread phenomenon, can spread to unmanaged edge habitats, such as shores of lakes and ponds, or borders between forests and open wetlands or forests and natural clearings or barrens such as rock outcroppings. Although management pressures change the nature of edge habitats, there is some overlap in the species that can occur in both. The widespread dominance of degraded anthropogenic edge habitats dominated by invasive species creates a reservoir of invasive plants such that there are more seed sources for these plants, relative to the native species that would colonize natural edge habitats.
How you can help: tips and tricks
The most important thing you can do to combat the bias against tall plants is to become aware of it, reflect on it, and move beyond it on your own. There are also some tricks to add to your toolbox that can lead to a "best of both worlds" for preserving populations of tall plants on sites where their height poses practical problems.Utilize topography
One of the best ways to utilize tall plants is in small depressions on an otherwise dry, upland site or site with poorer soils. These depressions accumulate both moisture and litter, forming islands of richer, moister soil in drier, nutrient-poor surroundings. Their natural equilibrium is thus to support taller plants. You can utilize this gradient of conditions in a flower bed, planting tall plants that will stay put in the central dip, and shorter, more drought-tolerant plants around the outside of the bed. The lower height in the dip will also level out the plant heights somewhat.Use stakes, trellises or other support
Although these practices are more labor-intensive and not practical on a large scale, gardeners who wish to grow tall plants in a small space can use their own structures such as a trellis or stakes to support tall plants. These supports mimic the natural support these plants would have from other vegetation, and they can keep the plants out of paths or roads.Growing tall plants but keeping them short: the "Chelsea chop"
If you want to grow species of plants that naturally would grow tall, on a site rich enough to support such height, but you want to keep them shorter for practical reasons, a useful trick is the "Chelsea chop". The Chelsea chop refers to the practice of cutting perennials or biennials fullly or partway to the ground mid-growing-season, typically in late spring. It is named for the Chelsea Flower Show in London, which is typically held in late May; the name originated in reminding people who were aware of the dates of the show, when to cut back their plants, but it has persisted in usage even in North America where few people are aware of the dates of the actual show. Some people remember the timing as being between mother's and father's day, although this timing allows for it to be a few weeks later.Most perennials and biennials are resilient to top-kill, and this is especially true of plants that naturally grow taller, as the lush environments where these plants thrive tend to attract heavy herbivore browsing, such as White-tailed deer in the gaps and edges in the Eastern Temperate Forests, and, historically, bison in the Great Plains. If you cut off these plants, they will simply regrow. Their maximum height will be reduced, but they will usually still flower.
 Plants native to the tallgrass prairie are adapted to top-kill during the growing season, through disturbances such as feeding from bison which historically inhabited the area. Photo © US Forest Service, Public Domain, Source.
Plants native to the tallgrass prairie are adapted to top-kill during the growing season, through disturbances such as feeding from bison which historically inhabited the area. Photo © US Forest Service, Public Domain, Source.The timing of the chop is important: it must be late enough to remove growth from a significant portion of the time plants were extending their height, but early enough not to affect flowering. Early bloomers, such as foxglove beardtongue (Penstemon digitalis) in the east and many other Penstemon species, will have their flowering negatively affected by the chop if too much is removed. But most taller plants bloom later, either in mid-summer at the earliest, or in late summer to fall. A full top-kill in late May will give these plants ample time to regrow, flower profusely, and produce seeds.
You can use the Chelsea chop to grow otherwise tall plants in areas such as next to paths and roads where flopping plants are a liability. You can also use it selectively on only a few isolated tall plants if you want to allow other, shorter plants to remain competitive with them, to keep a balance that can increase biodiversity. And you can use it for purely aesthetic reasons.
Park and land managers and road crews can also use the same general principle, mowing or bushwhacking a narrow strip next to paths and roads once a season, around the same time, so as to reduce vegetation flopping into the pavement, while allowing some taller plants to still grow up during the peak of the growing season. In much of North America, where sudden downpours from thunderstorms are more common in summer, this mow timing also protects soil by keeping vegetation lush during the the time when erosion risk is highest.
The optimal timing and cut length vary by vegetation type, site, and region. But the optimal timing usually falls somewhere in late spring, and plants can be cut back either to the ground, only a small distance, or anywhere in between.
In Summary
The bias against tall plants has a myriad of negative effects on North America's ecosystems. Its effects include the introduction, breeding, and spread of invasive plants originating as turfgrass, groundcovers, and lawn weeds. The breeding of shortened cultivars of native plants may weaken populations of those species, and at a bare minimum, the direct offspring of these plants are less competitive in the wild, reducing the benefit of growing them in gardens. The exclusion of tall plants from ecological restoration projects can lead to the failure of those projects and their becoming overrun by invasives. And lastly, the targeting of tall plants in the management of edge habitats can degrade these habitats and lead to the establishment of invasive plants both in the interiors behind the managed spaces, and in the disturbed areas along the edge itself. These edges then become networks through which the invasive plants spread throughout whole regions.You can help address these issues first by becoming aware of them, and then by talking about them with others. If you are in a decisionmaking position in any organization or project where these issues are relevant, use your knowledge to make more informed choices. There are also numerous techniques that can help you to use tall plants even on sites where their height may pose problems. Utilizing topography and planting taller plants in dips, using support, and cutting plants back to the ground in late spring, a.k.a. the "Chelsea chop", are three techniques to incorporate tall plants into managed landscapers where flopping plants are a liability.
A Focus on Goldenrods (Solidago sp.)
July 23rd, 2025 by Alex Zorach
When we launched our site and began prioritizing which plant articles and ID guides to complete, the initial focus was on trees, which made sense because trees are the largest plants in an ecosystem, there are relatively fewer tree species, and there were more sources writing about them.However, herbaceous plants (plants that do not grow woody parts) are also important to ecosystems, and there are some regions, such as the Great Plains, where they are more dominant than woody plants. And arguably one of the most important genera of herbaceous plants in north America is the goldenrods (Solidago sp.)
 This patch of Nevada goldenrod (Solidago spectabilis) is growing at Inyo National Forest in California. Although best-known as species of grasslands such as meadows, prairies, and savannas, goldenrods occur in habitats as diverse as deserts, forests, and wetlands. This species prefers locally-wet sites such as seeps, springs, and streambanks in deserts, mountain valleys, and alkali flats. Photo © mel letterman, CC BY 4.0, Source.
This patch of Nevada goldenrod (Solidago spectabilis) is growing at Inyo National Forest in California. Although best-known as species of grasslands such as meadows, prairies, and savannas, goldenrods occur in habitats as diverse as deserts, forests, and wetlands. This species prefers locally-wet sites such as seeps, springs, and streambanks in deserts, mountain valleys, and alkali flats. Photo © mel letterman, CC BY 4.0, Source.Including hybrids, BONAP recognizes a whopping 100 species of goldenrod in North America, all of which are native. We currently list 77, as we have not yet listed all hybrids. There are even some species that have been split from BONAP's 2014 listing, so the final count may number higher than 100.
Lately we have begun the difficult task of building ID guides for the goldenrods of North America. There are several reasons we have chosen to prioritize these plants, and I hope this post can highlight both the goldenrods and our reasons for prioritizing them.
Goldenrods as Keystone Species
Goldenrods are often hailed as keystone species, meaning that they have disproportionately large effects on their environment relative to their abundance. The importance of goldenrods in their environment is not limited to any one factor, so we will explore several.High Biodiversity in North America
Although goldenrods are found on several continents, including Europe, Asia, South America, and a small region in northwest Africa, North America is the global center for goldenrod diversity; only 4 are found in South America and 6-10 in Europe, depending on how you classify them.The diversity among goldenrod species enables them to thrive in surprisingly different habitats. People mostly think of goldenrods occurring in disturbed, open, sunny habitats, but there are numerous species that are found in shade, and they have partitioned the niches even in these habitats. For example, in the east, zigzag goldenrod (Solidago flexicaulis) is found in moist, rich forests, often on north- or east-facing slopes. Blue-stemmed goldenrod (Solidago caesia) also occurs on woods and in slopes, but usually on drier sites, often with some clay and/or rock in the soil. Another woodland goldenrod, elmleaf goldenrod (Solidago ulmifolia), is usually found on dry uplands with calcareous soils.
 Zigzag goldenrod (Solidago flexicaulis) is often seen as atypical among goldenrods in that it grows in shade, typically in forests, and its inflorescence shape is also different, blooming out of the upper leaf axils, not just a "rod" shape at the top of the stem. But there are actually numerous goldenrods that grow in forests, as well as others that have this axillary bloom pattern. Photo © Tom Scavo, CC BY 4.0, Source.
Zigzag goldenrod (Solidago flexicaulis) is often seen as atypical among goldenrods in that it grows in shade, typically in forests, and its inflorescence shape is also different, blooming out of the upper leaf axils, not just a "rod" shape at the top of the stem. But there are actually numerous goldenrods that grow in forests, as well as others that have this axillary bloom pattern. Photo © Tom Scavo, CC BY 4.0, Source.Seaside goldenrod (Solidago sempervirens) has adapted itself to coastal sand dunes with their coarse sandy soils and salt spray. In the north, Bog goldenrod (Solidago uliginosa), as its name suggest, grows in bogs. In the southwest, Wright's goldenrod (Solidago wrightii) grows on dry hillsides and Juniper-Pinyon woods. And these are all what one might think of as "atypical" goldenrods. There is even a goldenrod that isn't yellow: white goldenrod (Solidago bicolor).
The species richness in goldenrods is directly related to their abundance and wide distribution in different habitats, which is in turn related to their support of insects, as insects tend to evolve to eat plants that are abundant.
Support for Insects
It is hard to write a section about how many insects goldenrods support, as there is so much to say. Goldenrods are best known for their showy flowers and support for pollinators. Their inflorescences are large, with long bloom periods in which new flowers open as others are pollinated. The nectar tubes tend to be short and accessible, with spots for larger insects to perch. So the flowers tend to attract small and large pollinators alike, both short-tongued and long-tonged bees, as well as wasps, flies, and lepidoptera (butterflies, skippers, and moths), as well as less-well-known pollinators such as beetles, ants, and even the occasional true bug. Each of these groupings of insects has numerous different species which visit goldenrod flowers, and in the cases of the bees, wasps, and flies, the list is quite long. This photo shows a blue-winged scoliid wasp (Scolia dubia) feeding on Goldenrod. Adults of this wasp are attracted to goldenrod nectar, but in their larval form they eat beetle larvae. The presence of goldenrods thus exerts an indirect effect on controlling the populations of herbivorous beetles. Goldenrods belong to hundreds of such ecological relationships, which is one reason why they are considered keystone species. Photo © Christopher Volker, CC BY 4.0, Source.
This photo shows a blue-winged scoliid wasp (Scolia dubia) feeding on Goldenrod. Adults of this wasp are attracted to goldenrod nectar, but in their larval form they eat beetle larvae. The presence of goldenrods thus exerts an indirect effect on controlling the populations of herbivorous beetles. Goldenrods belong to hundreds of such ecological relationships, which is one reason why they are considered keystone species. Photo © Christopher Volker, CC BY 4.0, Source.But pollinators are only a small portion of the insects supported by goldenrods. Numerous insects feed destructively on the various parts of the plants, including the leaves, stems, roots, and even other portions of the flowerheads. These herbivorous insects include both generalists, and a large number of specialists that use goldenrods as their preferred food source, and in a few cases, their only food source. Numerous lepidoptera eat goldenrods in their larval form; a lot of sources cite figures saying 100 or more species, but data from the UK Hosts Database, which is no longer publicly available but which we have archived and used for our internal analyses, shows an even higher count of 143 Lepidoptera species recorded eating goldenrods in North America.
So each goldenrod plant is teeming with insect life. The abundance of insects attracts predators and parasites. Both insect predators and other invertebrate predators such as spiders thrive on goldenrods. There are even specialist insects that prefer host or prey insects that themselves specialize on goldenrods. A whole miniature ecosystem is found within a single plant. And all these organisms then become food for larger animals such as birds.
High Biomass
Number of species alone is not the only measure of the importance of a particular genus; it is also relevant how common and dominant in their ecosystems particular plants are. In the case of goldenrods, many of them are both abundant and dominant in many ecosystems. One of the most dominant species, tall goldenrod (Solidago altissima), ranges across most of North America, and frequently forms clonal colonies with 40 or more stems each. Other aggressive species include canada goldenrod (Solidago canadensis) in the northeast, giant goldenrod (Solidago gigantea) in floodplains, wrinkleleaf goldenrod (Solidago rugosa) in the east, western goldenrod (Solidago lepida) in the west and north, and Missouri goldenrod (Solidago missouriensis) which is more dominant in the great plains. This list is hardly exhaustive. Tall goldenrod (Solidago altissima) is often a dominant component of the ecosystems where it occurs. It thus contributes both a high amount of total biomass and structure, to the ecosystem. Without large goldenrods, many ecosystems would be very different and much poorer as a result. Photo © botany08, CC BY 4.0, Source.
Tall goldenrod (Solidago altissima) is often a dominant component of the ecosystems where it occurs. It thus contributes both a high amount of total biomass and structure, to the ecosystem. Without large goldenrods, many ecosystems would be very different and much poorer as a result. Photo © botany08, CC BY 4.0, Source.Even "smaller" goldenrods are often robust relative to the ecosystems they occur in. For example, sweet goldenrod (Solidago odora) is often one of the larger perennials in the acidic, nutrient-poor soils it grows in.
The large biomass of these species amplifies their benefit to the food web, such that they support not only diversity, but also large numbers of insects. The volume of insects then supports larger animals such as nesting birds. But the contributions to the food web are only one among many ecosystem services provided by goldenrods.
The physical robustness of goldenrods, especially the taller species, makes them valuable as cover for animals, and structure that other plants can use for support, not just including small vines but also other, floppier herbaceous plants that cannot support their own weight. The dense structure also reduces wind speeds and moderates temperature and humidity at lower levels in a thicket of plants.
The below-ground biomass, including the aggressive and extensive root systems of large colonies of goldenrods, also plays a key role in soil health. The aggressive roots help to stabilize soil and prevent erosion. The feeding of insects on roots creates burrowing action that aerates soil and cycles nutrients down to lower layers of soil. This process plays a key role in improving the soil structure and ultimately paving the way for succession to larger trees on some sites, such as when Solidago altissima colonizes sites with rich but compacted soil.
By investing so much energy in below-ground growth, goldenrods help make ecosystems resilient in the face of disturbances. When above-ground vegetation is cleared, such as by fire, flooding, deposition of sediment, heavy browsing by large mammalian herbivores, or anthropogenic disturbances such as mowing or weedwhacking, goldenrods just resprout, often after sending their rhizomes and/or stems into new locations to reconfigure in response to shifting ground. Goldenrods are critical for quickly revegetating disturbed sites, covering the ground with new foliage, and thus both reducing erosion, and moderating the ground-level temperature swings that often follow any disturbance that exposes the ground to sunlight. In the regions in which they are native, goldenrods can be seen as part of nature's healing mechanisms following a disturbance.
Maintenance of Open Habitats
Especially in the more humid parts of North America, such as the east and northwest, goldenrods play a key role in keeping open habitats open. Most of eastern North America has rich enough soil and high enough rainfall to support closed-canopy forests in the absence of disturbance that kills or removes trees. Following European colonization of North America, fire suppression, both intentional and unintentional (such as through building roads, which act as firebreaks) has led to the reduction of open habitats in areas of the east where land is left wild. A disproportionate number of open habitats are now managed, such as mowed lawn or cropland, not open habitat such as meadows, prairies, or savannas.Most goldenrods tend to grow on open sites such as meadows and prairies. But the relationship between goldenrods and open habitats is not merely one-directional: some goldenrods, including tall and Canada goldenrod, are allelopathic, meaning that they produce chemicals that inhibit the growth of other plants. This process delays, but cannot fully prevent, the colonization of open sites by trees. In slowing the growth of woody plants, these goldenrods protect open habitats, causing them to persist longer even in the absence of fire.
Resistance to Invasive Species
The combination of allelopathy and general robustness makes goldenrods a good buffer against a newer threat to ecosystems: invasive plants, plants that have been introduced across a major geographic divide, and threaten to disrupt ecosystems, often because native herbivores are not adapted to eat them. An invasive plant is generally going to have a much tougher time invading a habitat with goldenrods than one without. This colony of giant goldenrod (Solidago gigantea) under a power line clearance in Wadesboro, NC is competing successfully with numerous invasives, including Japanese honeysuckle (Lonicera japonica) and kudzu (Pueraria montana), two of the most aggressive ones. Plants like this both compete for light, and compete in the rootspace for water and nutrients. Photo © Becky Dill, Public Domain, Source.
This colony of giant goldenrod (Solidago gigantea) under a power line clearance in Wadesboro, NC is competing successfully with numerous invasives, including Japanese honeysuckle (Lonicera japonica) and kudzu (Pueraria montana), two of the most aggressive ones. Plants like this both compete for light, and compete in the rootspace for water and nutrients. Photo © Becky Dill, Public Domain, Source.I have also seen goldenrods competing, sometimes successfully, against some of the worst invasives, such as giant goldenrod pushing up through a patch of the Eurasian common reed (Phragmites australis), or tall goldenrod holding its own against common mugwort (Artemisia vulgaris). Sometimes goldenrods can even compete against invasives in unexpected, novel anthropogenic habitats, such as seaside goldenrod (Solidago sempervirens) seeding into patches of giant foxtail (Setaria faberi) in sandy and gravely soil along a railroad track.
The insects supported by goldenrods can also indirectly protect against invasion by other invasive insects. The insect community supported by goldenrods includes numerous generalist predators, such as praying mantises, spiders, and all manners of predatory bugs and beetles. Goldenrods also directly support larger animals, especially birds, which not only feed on the insect food but also directly on goldenrod seeds. Many of these predators will happily devour a random new insect that comes their way, thus making a habitat with goldenrods a lot more hostile towards a potential invasive insect.
Wrongly Blamed As Allergens
In the past, goldenrod was blamed for allergies, particularly seasonal "hay fever" which tends to strike in late summer into early fall, around when most goldenrods bloom. However, goldenrods are insect-pollinated and thus have heavy, sticky pollen that tends not to become airborne and people tend not to breathe in. Even people who react to goldenrod on a histamine allergy test will typically not experience many allergy symptoms caused by these flowers.The true culprit in "goldenrod" allergies are a close relative: ragweeds (Ambrosia sp.), which often grow together with goldenrods. Common ragweed (Ambrosia artemisiifolia) often grows together with tall goldenrod (Solidago altissima) in sunny, slightly dry habitats, and giant ragweed (Ambrosia trifida) often grows with giant goldenrod (Solidago gigantea) in moist bottomlands. Ragweeds have light, wind-borne pollen, and allergies to them are common and often quite unpleasant. But because ragweed's flowers are wind-pollinated, they are not showy, and as such, when people walk through an area full of ragweed, they often notice the showy goldenrod flowers and not the ragweed, and wrongly associate their allergy symptoms with the goldenrod.
 A typical sight in rich, moist bottomlands, giant ragweed (Ambrosia trifida) is growing together with giant goldenrod (Solidago gigantea) (lower-left and top-right corner). People may notice their allergies flare up when walking by such plant associations. Because the ragweed flowers (greenish conical structures) are easily overlooked, whereas the goldenrod flowers are visible and distinctive, many people form a mental association of goldenrods as causing their allergies. However, ragweed is usually the culprit. Photo © Eric Schmidt, CC BY 4.0, Source.
A typical sight in rich, moist bottomlands, giant ragweed (Ambrosia trifida) is growing together with giant goldenrod (Solidago gigantea) (lower-left and top-right corner). People may notice their allergies flare up when walking by such plant associations. Because the ragweed flowers (greenish conical structures) are easily overlooked, whereas the goldenrod flowers are visible and distinctive, many people form a mental association of goldenrods as causing their allergies. However, ragweed is usually the culprit. Photo © Eric Schmidt, CC BY 4.0, Source.A side note is that this pattern plays out in general: pollen allergies tend to be caused by tall, wind-pollinated plants, which tend to have less-showy flowers. Such plants include trees such as oak or mulberry, and various conifers, but not showy flowering trees such as cherry or magnolia. Similarly, they include plants such as grasses and nettle-family plants. Some genera of wind-pollinated plants, such as pellitory (Parietaria sp.) are important allergens globally, but the locally-native species, such as Pennsylvania pellitory (Parietaria pensylvanica), are so small that they rarely cause much trouble for people.
Utilizing Goldenrods
Goldenrods can be used both in gardening and landscaping, and in ecological restoration projects. The common complaint I hear about goldenrods is that they are "too aggressive" and "will take over your garden". While there is some truth in these remarks, at least for certain species, there are so many different goldenrods that there is almost always at least one or two species you can choose for a garden that will grow well, but without taking over or creating a problem. Many gardeners object to goldenrods as being too large and aggressive, but there are numerous goldenrods adapted to drier, harsher habitats that are smaller in size and less aggressive. Pictured here is gray goldenrod (Solidago nemoralis), a delicate plant that does not overwhelm other vegetation, and usually only grows where the soil is too dry and nutrient-poor for there to be much competition. Photo © Martyn Drabik-Hamshare, Public Domain, Source.
Many gardeners object to goldenrods as being too large and aggressive, but there are numerous goldenrods adapted to drier, harsher habitats that are smaller in size and less aggressive. Pictured here is gray goldenrod (Solidago nemoralis), a delicate plant that does not overwhelm other vegetation, and usually only grows where the soil is too dry and nutrient-poor for there to be much competition. Photo © Martyn Drabik-Hamshare, Public Domain, Source.Some of the goldenrods that prefer drier habitats with poorer soils make a better choice for gardens. One such example would be early goldenrod (Solidago juncea), and an example favoring even drier sites would be gray goldenrod (Solidago nemoralis). These species tend to grow slower and reach a smaller maximum size, and their greater drought tolerance can make them look good even in a dry year without watering. Their lower nutrient needs can eliminate the need for fertilizing. They make excellent choices for a bed where you add sand or gravel instead of mulch. They also can be a great choice for urban areas where the conditions are difficult due to reflected heat from pavement and buildings.
Even the more aggressive goldenrods can sometimes be utilized effectively in gardens. Tall and Canada goldenrods are showy, mindlessly easy to grow, and tend to shut out most competition, reducing the need for weeding. Giant goldenrod can also be used effectively in gardens when there is a bed that slopes down into a low, wet area in the back; in this scenario the giant goldenrod will thrive in the low area, pushing up through other plants, but will usually not move up into the drier part of the bed.
Where goldenrods usually shine the most, however, is in mass plantings in ecological restoration work. Giant goldenrod is ideal for riparian restorations, plantings to control erosion along streambanks and in floodplains. Tall goldenrod can be planted along roadsides and sunny edge habitats where soil is nutrient-rich but may be compacted or disturbed. Seaside goldenrod can even be useful for reclamation of former industrial sites, where it tolerates thin soils as well as the presence of salt and some minerals that would kill other plants. It also can thrive in roadside plantings due to its tolerance of drought and road salt.
Identification as Key to Understanding
The best way to understand plants and their habitat needs and ecological relationships is to observe them yourself, in the wild, in areas around you. To this end, it is helpful to know what you are looking at. As such we have prioritized completing ID guides, in many cases before we even complete the articles on each plant.We invite you to check out our currently completed ID guides on the Solidago genus, and to stay tuned for the completion of more guides.
Completed Solidago ID Guides
| Guide | Date Started↓ | Date Updated |
| Blue-Stemmed Goldenrod (Solidago caesia) vs. Mountain Decumbent Goldenrod (Solidago curtisii) | Mar 06, 2025 | Jul 20, 2025 |
| Tall Goldenrod (Solidago altissima) vs. Sweet Goldenrod (Solidago odora) | Mar 06, 2025 | Jul 20, 2025 |
| Early Goldenrod (Solidago juncea) vs. Sweet Goldenrod (Solidago odora) | Mar 05, 2025 | Jul 20, 2025 |
| Canada Goldenrod (Solidago canadensis) vs. Giant Goldenrod (Solidago gigantea) | Jan 16, 2025 | Jan 17, 2025 |
| Giant Goldenrod (Solidago gigantea) vs. Early Goldenrod (Solidago juncea) | Jan 12, 2025 | Feb 07, 2025 |
| Tall Goldenrod (Solidago altissima) vs. Giant Goldenrod (Solidago gigantea) | Dec 04, 2024 | Feb 08, 2025 |
| Tall Goldenrod (Solidago altissima) vs. Canada Goldenrod (Solidago canadensis) | Jul 20, 2023 | Feb 15, 2025 |
Under Construction Solidago ID Guides
| Guide | Date Started↓ | Date Updated |
| Early Goldenrod (Solidago juncea) vs. Gray Goldenrod (Solidago nemoralis) | Feb 27, 2025 | Feb 27, 2025 |
| Canada Goldenrod (Solidago canadensis) vs. Wrinkleleaf Goldenrod (Solidago rugosa) | Feb 05, 2025 | Feb 05, 2025 |
| Tall Goldenrod (Solidago altissima) vs. Wrinkleleaf Goldenrod (Solidago rugosa) | Jan 23, 2025 | Jan 28, 2025 |
| Blue-Stemmed Goldenrod (Solidago caesia) vs. Elmleaf Goldenrod (Solidago ulmifolia) | Jan 12, 2025 | Jan 12, 2025 |
| Wrinkleleaf Goldenrod (Solidago rugosa) vs. Elmleaf Goldenrod (Solidago ulmifolia) | Jan 12, 2025 | Jan 31, 2025 |
Thank you for your support!
We want to thank all our donors for supporting our ongoing work on this and all our other projects, such as the plant and ecoregion articles and maps. bplant.org remains a mostly-volunteer effort that currently provides only minimal compensation to me, and we donate a modest amount to iNaturalist which we use as the primary source for open-licensed images that make these ID guides possible. You can read more details on our finances page which is always accessible in the footer. Anyone wishing to donate can use the link in the header or footer, or you can get in touch with us about other ways to contribute.Archive of All Blogs
How To Identify Plants, May 7th, 2026
The Bias Against Tall Plants, November 11th, 2025
A Focus on Goldenrods (Solidago sp.), July 23rd, 2025
Disturbance and its Role in Plant Habitat Preferences, May 29th, 2025
What "Native" or "Introduced" Mean: Myths and Misconceptions, March 11th, 2025
Smarter & More Informative Search Results, January 13th, 2025
The Effect of the 2024 US Election on Plant Biodiversity and bplant.org, October 30th, 2024
The Problems With Nursery-Bought Plants, And The Solutions, October 8th, 2024
More Databases Linked & Search Improvements for Scientific Names, July 9th, 2024
Choosing The Best Common Names For Plants: Challenges & Solutions, April 19th, 2024
Range Map & Taxonomic Update Progress, January 31st, 2024
Giving Thanks To Everyone We Rely On, November 22nd, 2023
Thinking More Deeply About Habitat, April 5th, 2023
2022 Year-End Summary: Successes & New Goals, February 15th, 2023
New Databases Linked: Flora of North America & NatureServe Explorer, November 11th, 2022
All Range Maps 2nd Generation, Taxonomic Updates, & Fundraising Goal Met!, September 29th, 2022
More Range Map Improvements, POWO Interlinking, And Notes Fields, June 7th, 2022
Ecoregion-Based Plant Lists and Search, March 30th, 2022
Progress Updates on Range Maps and More, February 10th, 2022
The Vision for bplant.org, December 9th, 2021
New Server: Software & Hardware, August 30th, 2021
More & Improved Plant Range Maps, July 19th, 2021
A Control Section for Invasive Plants, April 15th, 2021
Progress Bars & State Ecoregion Legends, March 11th, 2021
Our 2020 Achievements, February 9th, 2021
Interlinking Databases for Plant Research, November 11th, 2020
A New Homepage, Highlighting Our Articles, July 29th, 2020
A False Recovery, But North Carolina's Ecoregions are Complete!, June 9th, 2020
We're Back After COVID-19 Setbacks, April 3rd, 2020
Help Us Find Ecoregion Photos, February 27th, 2020
What We Achieved in 2019, December 30th, 2019
Plant Comparison and ID Guides, October 30th, 2019
We Are Now Accepting Donations, October 14th, 2019
US State Ecoregion Maps, New Footer, Ecoregion Article Progress, and References, September 19th, 2019
Tentative Range Maps of Native Plants, August 12th, 2019
Ecoregion Locator and Interactive Maps, July 10th, 2019
Using Ecoregions Over Political Boundaries, May 13th, 2019
How We Handle Wild vs Cultivated Plants, April 16th, 2019
A Blog To Keep People Updated On Our Progress, April 8th, 2019
Sign Up
Want to get notified of our progress? Sign up for the bplant.org interest list!